Competing for Consciousness:
A Darwinian Mechanism at an
Appropriate Level of Explanation
William H. Calvin
University of Washington
Department of Psychiatry and Behavioral Sciences
Seattle WA 98195-1800 USA
WCalvin@U.Washington.edu
www.WilliamCalvin.com
Abstract. Treating consciousness as awareness or attention
greatly underestimates it, ignoring the temporary levels of
organization associated with higher intellectual function
(syntax, planning, logic, music). The tasks that require
consciousness tend to be the ones that demand a lot of
resources. Routine tasks can be handled on the back burner but
dealing with ambiguity, groping around offline, generating
creative choices, and performing precision movements may
temporarily require substantial allocations of neocortex. Here I
will attempt to clarify the appropriate levels of explanation
(ranging from quantum aspects to association cortex dynamics)
and then propose a specific mechanism (consciousness as the
current winner of Darwinian copying competitions in cerebral
cortex) that seems capable of encompassing the higher
intellectual function aspects of consciousness as well as some of
the attentional aspects. It includes features such as a coding
space appropriate for analogies and a supervisory Darwinian
process that can bias the operation of other Darwinian
processes.
Francis Crick likes to observe that people once worried
about the boundary between the living and the nonliving.
Today, the boundary seems meaningless; we instead talk
about all the varied aspects of molecular biology. Today's
brain researchers think it likely that much of the present
scientific and philosophical concern about consciousness will
soon become equally obsolete, that we will simply come to talk
of the various physiological processes involved with attention
and creative problem-solving.
 This is not to say that consciousness will disappear as a
useful word in our vocabularies. Nor will the philosophers'
cautions about classical mistakes cease to be relevant. Just as
"living" has continued to be a useful shortcut, so will
"conscious" and "mind." Even those of us who come to
understand all the contributing physiological processes will
still need the C word, not only for communicating with those
outside the field but even for thinking about the subject.
This is not to say that consciousness will disappear as a
useful word in our vocabularies. Nor will the philosophers'
cautions about classical mistakes cease to be relevant. Just as
"living" has continued to be a useful shortcut, so will
"conscious" and "mind." Even those of us who come to
understand all the contributing physiological processes will
still need the C word, not only for communicating with those
outside the field but even for thinking about the subject.
So let me attempt a neurophysiologist's introduction to
the subject, one that I hope will remain useful as we build our
knowledge about what Dan Dennett called "the last surviving
mystery." A mystery, Dennett said, "is a phenomenon that
people don't know how to think about -- yet." Here I will
attempt to clarify the appropriate levels of explanation and
then propose a candidate mechanism, a Darwin Machine
(Calvin 1987, 1996) that seems capable of encompassing the
higher intellectual function aspects of consciousness as well as
some of the attentional aspects. [Portions of this article are adapted from my two 1996 books,
How Brains Think (Basic Books, New York) and The Cerebral Code
(MIT Press, Cambridge
MA).].
Consciousness is the tip of the iceberg, in the sense that
many other things are going on in the brain at the same time,
hidden from view. Indeed, we are incapable of reporting on
most of what our brain does for us. I cannot report on my
blood pressure without a lot of external instrumentation, even
though my brain measures and regulates my systolic and
diastolic pressures. It is simply not accessible for verbal
reporting.
For the brain, we need to span multiple levels of
mechanism -- from synapses to cells to circuits to modules,
and more. And also span multiple levels of phenomenological
explanation -- such as attributes, objects, categories, analogies,
and metaphors. Layers of middlemen are familiar from
everyday economics, and we expect to see many layers of
representation standing between our consciousness and the
real world. As Derek Bickerton noted:
[T]he more consciousness one has, the more layers of
processing divide one from the world. . . . Progressive
distancing from the external world is simply the price
that is paid for knowing anything about the world at all.
The deeper and broader [our] consciousness of the world
becomes, the more complex the layers of processing
necessary to obtain that consciousness.
There are subconscious trains of thought, however, and
they vie for "attention." The layer immediately underlying
consciousness might well involve a mechanism for competition.
Though the obvious analogy is to the television viewer
who surfs the channels (and our nighttime dreams often seem
like switching between soap operas in progress), there need not
be a central place where choices are viewed. The "best"
channel need only temporarily win out over the others in the
battle for access to output pathways such as speech and other
body movements. Soon, another channel comes to dominate
and we speak of "our attention shifting" -- but there need not
be an agent which makes the decision or performs the action,
not any more than ice needs an agent to help it melt.
Output need not involve overt movements: silent
speech is an important way in which we reason, generate
alternatives, and choose amongst them. The mental rotation of
complex objects illustrates a nonlanguage mode of conscious
thought. But, to a neurophysiologist, there is nothing in this
overview that demands a central place. The "center of
consciousness" could, instead, shift from moment to moment:
from language to nonlanguage areas, from frontal to parietal
lobe, from left to right hemisphere, and maybe even from
cortical to subcortical structures -- anywhere, I suspect, with
the potential for generating novel patterns of movement. And,
of course, it seems possible to do several things at once, just as
one can sew, or listen to the radio, while watching television.
Not everything we do has to pass through the current center of
consciousness.
Karl Popper noted that posing alternatives seemed especially
“conscious”:
Much of our purposeful behaviour (and presumably of
the purposeful behaviour of animals) happens without
the intervention of consciousness.... Problems that can
be solved by routine do not need consciousness. [The
biological achievements that are helped by
consciousness are the solution of problems of a
non-routine kind.] But the role of consciousness is
perhaps clearest where an aim or purpose... can be
achieved by alternate means, and when two or more
means are tried out, after deliberation.
While I think that Popper is on to something here, I'd state it
more generally: the tasks that require consciousness tend to be
the ones that demand a lot of resources. Routine tasks can be
handled on the back burner but dealing with ambiguity,
groping around, generating creative choices, and performing
precision movements may temporarily require substantial
allocations of neocortex.
Consciousness and Levels of Explanation
What constitutes an explanation? To explain consciousness
in any serious way, we must avoid using mathematical
concepts to dazzle rather than enlighten. And we must watch
out for "proofs by want of imagination," as when we conclude,
out of arrogance or impatience, that there are no other
alternatives to the answers we have found. When it comes to
the brain, in particular, we must be careful to pitch our theories
at the right level of mechanistic explanation.
Brain function involves at least a dozen levels, and
finding appropriate levels for aspects of consciousness has not
been easy. Everyone tends to extrapolate their favorite subject
of study and proclaim its relevance to consciousness. Since I'm
about to do the same thing myself, indulge me in a few words
about levels, particularly as they relate to emergent properties
and those "changes of state," for I have two major points to
make in this short summary of (and borrowing from) my
longer works, How Brains Think and The Cerebral Code. One is a
proposal for a specific consciousness mechanism (as the current
winner of Darwinian copying competitions in association
cortex) and the other is an argument about what's a correct
level at which to seek a consciousness mechanism. I hope both
are correct, but the level analysis ought to be more generally
useful; it's certainly intended as a critique of much that has
been written on the subject.
What's missing from most discussions of consciousness
is, surprisingly, the whole concept that there are levels of
explanation. Douglas Hofstadter (1985) gives a nice example of
levels when he points out that the cause of a traffic jam is not to
be found within a single car or its elements. Traffic jams are an
example of self-organization, more easily recognized when
stop-and-go achieves an extreme form of quasi-stability -- the
crystallization known as gridlock. An occasional traffic jam
may be due to component failure, but faulty spark plugs aren't
a very illuminating level of analysis-- not when compared to
merging traffic, comfortable car spacing, driver reaction times,
traffic signal settings, and the failure of drivers to accelerate for
a hill or high-rise bridge.
The more elementary levels of explanation are largely irrelevant to
traffic jams. Such decoupling was emphasized by the physicist Heinz
Pagels (1988), who
noted:
“Causal decoupling” between the levels of
the world implies that to understand the material basis
of certain rules I must go to the next level down; but the
rules can be applied with confidence without any
reference to the more basic level. Interestingly, the
division of natural sciences reflects this causal
decoupling. Nuclear physics, atomic physics, chemistry,
molecular biology, biochemistry, and genetics are each
independent disciplines valid in their own right, a
consequence of the causal decoupling between them....
Such a series of “causal decouplings” may
be extraordinarily complex, intricate beyond our current
imaginings. Yet finally what we may arrive at is a
theory of the mind and consciousness — a mind so
decoupled from its material support systems that it
seems to be independent of them — and
“forgot” how we got to it.... The biological
phenomenon of a self-reflexive consciousness is simply
the last of a long and complex series of “causal
decouplings” from the world of matter.
Stratified Stability and Emergents
Closely related is the notion of emergent properties: traffic
jams and crystals emerge from combinations, and we expect
emergence to play a large role in the transient levels of
organization involved with higher intellectual function
(language, planning, games, etc.). In our search for a level
corresponding to consciousness, it is well to recall that levels
arise from what Jacob Bronowski (1973) called stratified stability:
Nature works by steps. The atoms form molecules, the
molecules form bases, the bases direct the formation of
amino acids, the amino acids form proteins, and proteins
work in cells. The cells make up first of all the simple
animals, and then the sophisticated ones, climbing step
by step. The stable units that compose one level or
stratum are the raw material for random encounters
which produce higher configurations, some of which
will chance to be stable.... Evolution is the climbing of a
ladder from simple to complex by steps, each of which
is stable in itself.
The tumult of random combinations occasionally produces a
new form of organization. Some forms, such as the hexagonal
cells that appear in the cooking porridge if you forget to stir it,
are ephemeral (as, indeed, are the contents of our
consciousness). Other forms may have a "ratchet" that
prevents backsliding once some new order is achieved. While
crystals are the best known of these quasi-stable forms,
molecular conformations are another, and it is even possible
that there are quasi-stable forms at intermediate levels -- such
as the microtubule quantum states where the consciousness
physicists would like the action to be:
“Accordingly, the neuron level of description that
provides the currently fashionable picture of the brain
and mind is a mere shadow of the deeper level of
cytoskeletal action — and it is at this deeper level
where we must seek the physical basis of mind!”
Roger Penrose, Shadows of the Mind, 1994
While I hope that quantum field effects on consciousness
exist (as you will presently see, when I get to technological
possibilities), there are a number of problems with such
proposals, not the least of which is the scanty evidence relating
quantum fields to cortical synchrony, or relating synchrony to
perceptual binding, or relating binding to attention, or relating
attention to the much broader range of phenomena associated
with consciousness (the word has many connotations, and they
surely don't all share the same mechanism).
To limit myself to the initial leap from QM (my abbreviation for all
the “quantums” reviewed by Jibu and Yasue 1995) to
synchrony, one should always be suspicious of any mechanism that
claims to tunnel through a dozen levels of organization to produce a
striking effect. As I explained in How Brains
Think:
[A] more appropriate level of inquiry into consciousness
is probably at a level of organization immediately
subjacent to that of perception and planning: likely (in
my view) cerebral-cortex circuitry and dynamic self-organization involving firing patterns within a constantly
shifting quiltwork of postage-stamp-sized cortical
regions. Consciousness, in any of its varied
connotations, certainly isn't located down in the
basement of chemistry or the subbasement of physics.
This attempt to leap, in a single bound, from the
subbasement of quantum mechanics to the penthouse of
consciousness is what I call the Janitor's Dream.
Quantum mechanics is probably essential to consciousness in
about the same way as crystals were once essential to radios, or
spark plugs are still essential to traffic jams. Necessary, but not
sufficient. Interesting in its own right, but a subject related
only distantly to our mental lives -- and not to be confused with
the temporary levels of organization associated with higher
intellectual function.
Darwinian Bootstrapping of Quality
An existence proof for a good alternative is potentially far
more persuasive than the best critique of quantum-mechanical
spirits and little-man-inside reasoning, so let me shift gears
here. If we take consciousness to be important for Popper's
"solution of problems of the non-routine kind," then shaping
up a good-to-excellent ("quality") course of action in thought is a key aspect of
consciousness, one that goes well beyond mere awareness or
shifting attention.
We no longer have to take it on faith that there are
mechanisms capable of recursively bootstrapping random
novelties into something of quality. For the last 160 years,
there has been an existence proof, the Darwinian process
(Calvin 1996b, 1997). The way in which quality is achieved
using this process has long occupied the best minds in
evolutionary biology (see, for example, Maynard Smith and
Szathmáry 1995). And the slow evolution of species, on the
time scale of millennia, is no longer the only example: the
immune response is now known to be another Darwinian
process, operating on the time scale of days to weeks as a better
and better antibody is shaped up in response to the challenge
of a novel antigen. For decades, computer science has used a
solution-finding procedure, called the genetic algorithm
(Holland 1992), that mimics an expanded version of the
Darwinian process on a time scale limited only by computer
size and speed.
It would be surprising if the brain did not make some
use of this fundamental principle for bootstrapping quality.
Can this same well-known process operate quickly enough in
the brain, on the time scale of thought and action? Can it
account for much of what we call "consciousness"? I
undertook to answer such questions in one of my 1996 books,
The Cerebral Code, which analyzes the recurrent excitatory
circuitry of mammalian neocortex. I showed that this
widespread wiring principle was capable of running the six
essential features of the classical Darwinian process:
a pattern (spatiotemporal firing pattern of a
Hebbian cell-assembly, in this case) that copies
with occasional variation, where populations of
the variants compete for a limited work space,
their relative success biased by a multi faceted
environment (both memorized and real-time, in
this case), and with further variations centered on
the more successful of the current generation
(Darwin's inheritance principle).
This full-fledged Darwinian process is what is associated with
the recursive shaping up of quality; it should not be confused
with mere selective survival of a single pattern and other
"sparse sets" that utilize only a few of the "six essentials"
(Calvin 1997).
The best way of demonstrating what I consider an
appropriate level of explanation, that of circuits involving
stamp-to-postcard-sized areas of cerebral cortex (we have
enough neocortex to fill four sheets of typing paper), is to take
the reader through an example. It's not the only possible
example, just the one due to Darwin, but I expect that
competing theories will also come to occupy the same level of
explanation, that of emergent properties in the shifting
dynamics of substantial areas of cortex. After I explain enough
of the circuitry so that the reader can imagine how a Darwinian
process could operate in neocortex, I will briefly return to
Quantum Consciousness, trains of thought, and the
explanatory coverage needed for a theory of mind.
The Brain's Darwinian Circuitry
The cortical circuitry that makes a full-fledged Darwinian
process possible is not an obscure feature known only to a few
neuroanatomists: it is easily the most prominent wiring
principle seen in cerebral cortex, that of the patterned recurrent
excitatory connections between neighboring pyramidal
neurons in the top layers of neocortex. It has just taken a while
to realize one of the implications of it, an emergent property of
the circuit not possessed by any of the individual elements:
synchronized triangular arrays of pyramidal neurons, with
nodes about 0.5 mm apart, is what you expect to observe, some
of the time.
Each pyramidal neuron has an axon that branches nearly
10,000 times. Some travel through the white matter but most of
the branches never leave the cortical layers, terminating in a
synapse within a millimeter or so. The axon travels sideways
to excite other cortical neurons, mostly other pyramidal
neurons. The deep-layer (V and VI) pyramidal neurons have
such sideways axons that remain within the cortical layers,
some terminating nearby and others more distantly.
It's the wiring seen (e.g., Lund et al 1993) in the
branching of the axon of the superficial-layer pyramidal
neurons (layers II and III), however, that is so striking. Their
terminations are patterned: their axons are like express trains
that skip a long series of intermediate stops, concentrating their
synaptic outputs in zones about 0.5 mm apart. That's what
makes synchronized triangular arrays likely to form on
occasion.
Though the 0.5 mm spacing is similar to that of
macrocolumns (say, the orientation columns of visual cortex)
and though it may be related to helping organize such
macrocolumns during development (Calvin 1995), the
analogies to macrocolumns (as we currently know them) are
limited. What's intriguing is their relation to minicolumns
(orientation columns are the best known examples), which are
vertical columns of about a hundred neurons that are
organized around a dendritic bundle in the manner of stalks of
celery; the distance between neighboring bundles is about
0.023-0.031 mm (in monkey; twice that in cat: Peters and
Yilmaz 1993). The nearest outputs of a superficial pyramidal
neuron's axon are not 0.5 mm away but to immediate
neighbors. Cells within a minicolumn tend to share interests,
as in elongated visual stimuli of a particular orientation, and
this local recurrent excitation is one of the reasons.
 The longer axon branches tend to skip making synapses
for 0.4-0.6 mm, but the express-train analogy is too one-dimensional to capture the reality: axon branches go in many
radial directions. I tend to draw the result, looking down from
the top, as an annular ring; while no one cell's axon branches
will fill in the annulus, there are 39 superficial pyramidal
neurons in a given minicolumn doing similar jobs. So think of
the output pattern as something like a mal-focused flashlight
beam: a central minispot, then a bright ring in the periphery.
Actually, because many axons continue their express run for
another 0.5 mm, you need to imagine additional annuluses at
0.5 mm spacings, more like a bull's eye target with fading
shades of gray.
The longer axon branches tend to skip making synapses
for 0.4-0.6 mm, but the express-train analogy is too one-dimensional to capture the reality: axon branches go in many
radial directions. I tend to draw the result, looking down from
the top, as an annular ring; while no one cell's axon branches
will fill in the annulus, there are 39 superficial pyramidal
neurons in a given minicolumn doing similar jobs. So think of
the output pattern as something like a mal-focused flashlight
beam: a central minispot, then a bright ring in the periphery.
Actually, because many axons continue their express run for
another 0.5 mm, you need to imagine additional annuluses at
0.5 mm spacings, more like a bull's eye target with fading
shades of gray.
Synchrony on a Small Scale
One consequence of the express-train axon is that cells 0.5
mm apart will tend to talk to one another: they will recurrently
excite. While a chasing-their-tails loop is one possibility if
synaptic strengths are quite high, even weak synaptic strengths
have an important consequence: entrainment. Since 1665,
when the Dutch physicist Christiaan Huygens noticed that
pendulum clocks on the same shelf synchronized their ticks
within a half hour, much additional work has been done on
entrainment. A dramatic example from the Phillippines was
reported in Science by Hugh Smith in 1935:
Imagine a tree thirty-five to forty feet high, apparently
with a firefly on every leaf, and all the fireflies flashing
in perfect unison at a rate of about three times in two
seconds, the tree being in complete darkness between
flashes. Imagine a tenth of a mile of river front with an
unbroken line of mangrove trees with fireflies on every
leaf flashing in synchronization, the insects on the trees
at the ends of the line acting in perfect unison with those
between. Then, if one’s imagination is
sufficiently vivid, he may form some conception of this
amazing spectacle.
Even small tendencies to advance the next flash when
stimulated with light will suffice to create a "rush hour."
Furthermore, you usually do not see waves propagating
through such a population, except perhaps when the flashing is
just beginning or ending.
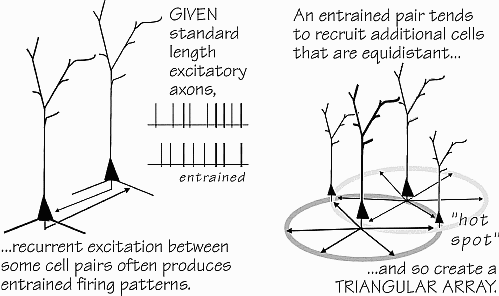
Relaxation oscillators like neurons and fireflies will get in sync much
more quickly than harmonic oscillators, and even weak interconnections will suffice (Somers and Kopell 1992). So, if several neurons
0.5 mm apart are firing for some reason (perhaps they both respond to
the color yellow), there is a good chance that they will get in sync
some of the time.
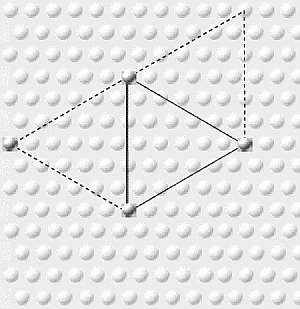 This has an interesting consequence: entraining
additional yellow neurons located 0.5 mm from each of the
original pair, to form an equilateral triangle of synchronized
superficial pyramidal neurons (as seen, looking down on the
cortex from above). Of course, yellow-2 and yellow-3 might
themselves entrain a fourth. What we have is a mechanism for
forming a triangular array of synchronized cells, one that can
extend its reach to wherever there are cells already firing, or
close to firing.
This has an interesting consequence: entraining
additional yellow neurons located 0.5 mm from each of the
original pair, to form an equilateral triangle of synchronized
superficial pyramidal neurons (as seen, looking down on the
cortex from above). Of course, yellow-2 and yellow-3 might
themselves entrain a fourth. What we have is a mechanism for
forming a triangular array of synchronized cells, one that can
extend its reach to wherever there are cells already firing, or
close to firing.
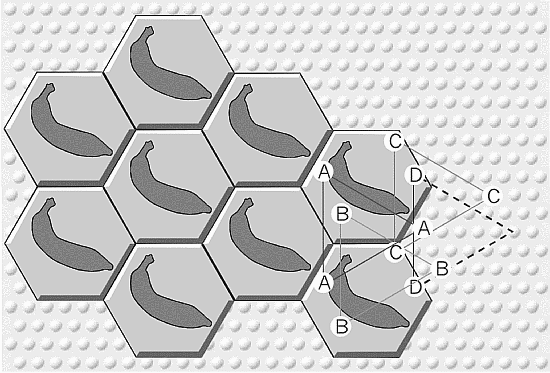
Suppose now that we have some other cells responding
to another attribute of a yellow banana, say its curvature. They
too can form their own triangular array. What is the largest
number of triangular arrays? Try to define an area containing
only one member of each triangular array. It turns out to be
hexagonal in shape (corresponding points in adjacent hexagons
are connected by equilateral triangles), 0.5 mm across, and
containing a few hundred minicolumns. Most potential arrays
will, of course, be silent; I tend to imagine fewer than a dozen
actively firing, but the silent ones are likely also important
(were they to become active, they might "fog" the characteristic
spatiotemporal firing pattern).
What we now have is a hexagonal mosaic, formed of the
active and silent triangular arrays. The adjacent hexagons are
nearly identical in their firing patterns. The curved array and
the yellow array need not be synchronized with one another (as
a common form of the binding theory assumes); indeed, they
are likely displaced in time. But the spatiotemporal firing
pattern within the hexagon is now the minimal one: it contains
one member of everything, but never a redundant one. The
microcircuitry within that 0.5 mm hexagon may resonate with
the spatiotemporal pattern. An imposed pattern might be
memorized via long-term potentiation and depression; indeed,
one can imagine reconstituting the spatiotemporal pattern if it
is halted, thanks to the lingering alteration of synapse strengths
from the imposed rhythms creating a new basin of attraction,
conforming any near-fit spatiotemporal patterns to the stored
standard.
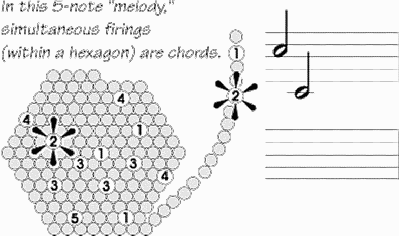
It's a minimal Hebbian cell-assembly, potentially capable
of recording the various features of an object or the details of a
movement program. And a hexagon's spatiotemporal firing
pattern is potentially a cerebral code, what represents an object
or idea. Such a pattern is like a little tune (map each of the
several hundred minicolumns to a note on a musical scale).
There will be a different tune characterizing Apple than the tune
for Banana.
The musical analogies also tell us that a hexagonal
mosaic is like a plainchant choir, singing in the lockstep of a
Gregorian chant. As the triangular arrays recruit followers on
the edges, additional hexagons are added to the mosaic.
Perhaps choosing between an apple and a banana for a snack is
a matter of how big their respective choirs are, that only large
choruses can gain access to the output pathways associated
with silent speech.
Dueling Choirs for Concept Competition
Now imagine dueling choirs, abutting hexagonal mosaics
singing different tunes, trying to recruit members at the
expense of the other. Along the battlefront, there are hexagons
that have both tunes superimposed, just as in a symphonic
work. If the combination resonates well with the local neural
network, we might speak of harmony, just as we do for the
major and minor scales. Borderline superpositions (as well as
the more extensive ones that can be created by long
corticocortical bundles) illustrate a powerful recombination
principle, a way of doing associative memories that can
represent relationships with the same 0.5 mm hexagonal code
space as used for objects.
 Another lesson of levels is that mechanisms that suffice
at one level may prove to be shaky foundations, that other
ways of doing the same thing are more extensible. Hexagonal
codes are a much better foundation for superstructures (such as
coding for analogies) than are the better-known associative
memory mechanisms at the level of synaptic mechanisms for
classical conditioning.
Another lesson of levels is that mechanisms that suffice
at one level may prove to be shaky foundations, that other
ways of doing the same thing are more extensible. Hexagonal
codes are a much better foundation for superstructures (such as
coding for analogies) than are the better-known associative
memory mechanisms at the level of synaptic mechanisms for
classical conditioning.
 This isn't the place for showing the many implications of
a cerebral coding scheme based on the spatiotemporal firing
pattern within one of the recurrent-excitation-defined
hexagons, a book-length project that I tackled in The Cerebral
Code. But with the notion of hexagonal mosaics that transiently
compete for space in association cortex, you can now
appreciate how a Darwinian process could operate in
association cortex via the spatiotemporal patterns copying
themselves sideways:
This isn't the place for showing the many implications of
a cerebral coding scheme based on the spatiotemporal firing
pattern within one of the recurrent-excitation-defined
hexagons, a book-length project that I tackled in The Cerebral
Code. But with the notion of hexagonal mosaics that transiently
compete for space in association cortex, you can now
appreciate how a Darwinian process could operate in
association cortex via the spatiotemporal patterns copying
themselves sideways:
Like the classical examples of a full-fledged
Darwinian process, there is a pattern that is copied
(indeed, what is reliably copied defined the
hexagonal-shaped spatiotemporal pattern),
variations occur (dropouts, off-focus nodes,
superpositions), populations of the variant
patterns compete for a work space, their relative
success is biased by a multifaceted environment
(current sensory as well as resonances with
memorized patterns), and the more successful of
the current patterns tend to produce more of the
next round of variants (Darwin's inheritance
principle is implemented because bigger mosaics
have more perimeter, and the perimeter is where
dropouts and off-focus nodes can escape the
standardization enforced by six surrounding
nodes all firing at the same time).
Unlike the generation times spanning days to decades of the
usual Darwinian examples, cortex operates on a time scale of
milliseconds to seconds, though its operations are biased by
memories that span far longer time scales. Within seconds to
minutes, neocortex ought to be capable of implementing all of
the classical means of accelerating the rate of evolution
(systematic recombination, parcellation, rapid "climate
change," and refilling empty niches).
A Web of Darwinian Competitions
Because of its distributed nature and corticocortical
connections between regions, cortex isn't limited to the
standard Darwinian productions. It might well utilize some
additional features, such as a supervisory Darwinian process
that can bias the operation of other Darwinian processes.
Yet it need not be some grand supervisor with even
more intelligence. Until something fancier is clearly indicated,
the default assumption ought to be that any regulatory process
is essentially stupid, perhaps only chaotic phenomena on a
grander or slower scale.
Indeed, there are some situations that might qualify for
such two-level interactive evolution, such as the orbital frontal
cortex role in monitoring progress on an agenda, a meta-sequence that seems to tick along on a different time scale than
individual thoughts and sentences. There's no requirement
that darwinian variations have to be truly random; a slow
darwinian process could bias or skew the general direction of
the variants of a faster darwinian process that deals with
lower-level matters, such as perception and movement on the
time scale of seconds. There could be a cascade or web of such
darwinian processes that operate on different levels in the
attributes-to-metaphors spectrum, or on different time scales in
the milliseconds-to-hours range.
The neocortical Darwin Machine theory seems to me to be
at the right level of explanation for consciousness; it's not
down in the synapse or cytoskeleton but up at the level of
dynamics involving tens of thousands of neurons, generating
the spatiotemporal patterns that are the precursors of
movement -- of behavior in the world outside the brain.
Composite cerebral codes, formed by superpositions and
shaped up by darwinian copying competitions, could explain
much of our subconscious mental lives. The codes themselves
are suitably arbitrary, just as a century of argument about
symbols has emphasized. Copying competitions suggest why
we humans can get away with many more novel behaviors
than other animals (we have offline evolution of nonstandard
movement plans judged for safety). It suggests how we can
engage in analogical reasoning (relationships themselves can
have codes that can compete). Because cerebral codes can be
formed from pieces, you can imagine a unicorn and form a
memory of it (resonances that can reactivate the spatiotemporal
code for unicorn). Best of all, a darwinian process provides a
machine for metaphor: you can code relationships between
relationships, and then shape them up into something of
quality.
None of these things naturally flow from Quantum
Consciousness. But let us not throw out the baby with the bath
water: there are interesting aspects of some of the quantum
field proposals.
The Quantums Revisited
Picking up on the notions of synchronization for binding the
dispersed aspects of the analysis on an object in the brain, some
have invoked a synchronizing influence of quantum fields,
suggesting a connection between QM and object recognition
(and, in the process, conflating perception with consciousness).
I hesitate to note that my synchronized triangular arrays for
pattern cloning show another possible application of the same
reasoning, extending the potential realm of QM beyond the
object-feature binding aspects of consciousness and into the
realm of higher intellectual function.
While it is possible that quantum-level phenomena
could influence some aspects of consciousness via encouraging
synchrony, extraordinary claims (QM mediating a key aspect
of consciousness) require extraordinary evidence. If the
consciousness physicists were serious about their proposal,
they would examine alternative ways of achieving synchrony
in the brain -- which are legion -- and explain why their
synchronizing explanation was preferable to simpler
explanations that are a dozen levels of organization closer to
higher intellectual function.
A caution, however: If I were to announce yet another
way of achieving cortical synchronization via some new
synaptic mechanism, everyone would yawn: given how many
ways there are of encouraging entrainment (even recurrent
inhibition can entrain; more generally, see Winfree 1967), the
name of the game is not possible mechanisms but probable
mechanisms. How strong is your candidate's synchronizing
influence? Do other known mechanisms seem to interdigitate
with it? Does it do anything that needs doing? (I'm convinced
of the need for a Darwinian process to bootstrap quality, but
some of us are not convinced that object attributes typically
need synchrony to bind them together after what-where
divergence.) The existence of other synchronization
mechanisms means that any QM effect could be easily
swamped, so proponents not only need to show a QM effect on
entrainment but to show the conditions when it can dominate
the others, and so bring Quantum Consciousness to the fore.
Such has led me to wonder if quantum-mechanical
proposals are not solutions in search of a problem. I'm not
convinced that brain science needs quantum-mechanical
mechanisms when we already have so many candidates at the
level of the emergent properties of circuits to accomplish the
same thing: induce entrainment among cortical neurons (never
mind getting from synchrony to binding, from binding to
attention, from attention to other aspects of consciousness).
But that's science -- and technology may well be another
matter, given my other role for entrainment, that of providing
the foundation for a hexagonal mosaic that can engage in a
Darwinian competition. Encouraging entrainment, or breaking
it up, via externally applied fields could provide an interface
with the Darwinian brain processes that shift the contents of
consciousness. If the postulated quantum effects prove strong
enough, and external fields can be focused sufficiently well to
elevate them to dominate other entrainment influences, such a
QM spotlight might make for an interesting technology. For
psychiatry, imagine breaking up obsessive patterns of thought
or suppressing hallucinations. For neurology, imagine
retraining cortical areas to take over a function lost elsewhere
to a tumor or stroke. For anesthesiology, imagine disrupting
those choirs that lead to memorizing unpleasant events.
Studying for exams with a quantum assist might aid in learning
new material; focusing the assist in other areas might aid in
recall.
The Stream of Consciousness
Resonances are better known these days as part of the rubric
of chaotic attractors. I imagine each hexagon's neural network
as supporting a number of characteristic spatiotemporal
patterns, just as spinal cord circuitry supports a number of
gaits, the particular spatiotemporal pattern that you get
depending on how you precondition the circuitry via the
facilitation from other imposed patterns. And that may have
something to say about the "stream of consciousness."
Manipulating the landscape of a basin of attraction is
reminiscent of William James's train of thought, that series of
mental states that preceded your current one, each one fading
into the background but overlain on its predecessors -- and all
capable of contributing to what connections you're likely to
make right now.
 Just imagine those various fading attractors as like that
Japanese technique of finely slicing some raw fish, then tilting
the block sideways (fallen dominos are another analogy, if you
are sashimi impaired). The bottom layer may be hardest to
reach but it goes back furthest. Stage-setting with multiple
layers of fading schemas may be handy for promoting creativity, getting the right layers of attractors in about the right
order and so adjusting their relative strengths. (The Sashimi
Theory of Creativity would, of course, be a suitably raw
successor to all those half-baked right-brain schemes).
Just imagine those various fading attractors as like that
Japanese technique of finely slicing some raw fish, then tilting
the block sideways (fallen dominos are another analogy, if you
are sashimi impaired). The bottom layer may be hardest to
reach but it goes back furthest. Stage-setting with multiple
layers of fading schemas may be handy for promoting creativity, getting the right layers of attractors in about the right
order and so adjusting their relative strengths. (The Sashimi
Theory of Creativity would, of course, be a suitably raw
successor to all those half-baked right-brain schemes).
But such histories can also be distracting, and we often
try to let them fade, try to avoid reexciting them with further
thinking. There are various mind-clearing techniques; Donald
Michael (personal communication 1995) suggests that forming
large quasi-stable hexagonal territories might be what
meditation with a mantra is all about, preempting the everyday
concerns that would otherwise partition the work space and
plate out new short-term attractors. By replacing it all with the
mantra's nonsense pattern, and holding it long enough for
temporary synaptic strength alterations to fade back to normal,
the meditator gets a fresh start (for things other than the
mantra!).
An ordinary mantra won't, of course, wipe the work
space clean: to prematurely erase those fading attractors, you'll
need a fancier mantra that disrupts instead. Short of fogging
with seizures, as in electroshock therapy, I don't know of any
such eraser schemas -- though one can imagine mental viruses
(Dawkins 1993) that might preempt entry into those fading
basins of attraction, more analogous to an obscuring coat of
paint than to a true eraser.
Higher Aspects of Consciousness
Once they finish with things as basic as perceptual transformations and memory phenomena, theories of brain function
must explain abstractions and associations as diverse as
categories, abstracts, schemas, scripts, syntax, and metaphor. If
we are to venture past the elementary notion of consciousness
as mere awareness or shifting attention, we are going to need
to account for:
- How items of our vocabularies are
represented.
- How memories are stored and recalled.
- How darwinian shaping-up takes place.
- How “new ideas” arise, perhaps as pattern variants or
superpositions.
Those four,
at least, have pretty much fallen out of our search for the darwinian
essentials. The rest are
harder:
- The existence of hallucinations and
dreams.
- Déjà vu experiences. Abnormally widespread cloning of an input pattern (perhaps
due to lack of competition) might produce the conscious experience
usually associated with strong memory resonances that allow
widespread cloning.
- Unreliable memories. Because the long-term synaptic connectivities can be modified by a new active pattern, this could often happen. Similarly, we’d like something appropriate for concrete thinking and the idée fixe.
- How the various connotations of a word such “comb” are linked, given that they’re likely stored in different cortical areas.
- The presence of specialized cortical regions that can also participate in nonspecialist tasks.
- The ability to hold a behavioral set, after selecting it from among possibilities. What might an agenda look like?
- Serial-order specialties for language and speculative planning, not to mention all the small muscle sequencing that a child needs to tie a
shoelace.
- Speed-of-thought correlates, mechanisms that could vary from time to time in the
same individual. During the transitions of manic-depressive illness, a
person can go from a fluidity of making connections and decisions to
a slow, labored train of thought that lingers too long and fails to make
obvious connections. And back
again.
- The impression of a narrator, juggling decisions, and speculating about
tomorrow. Any explanation needs to be consistent with the
neurological evidence that no partial cortical lesion abolishes the
“self.”
That's the kind of coverage needed for a useful theory of mind
(and this neocortical Darwin Machine enables predictions to be
made, all across this spectrum; Calvin 1996b). It may not have
to explain all of two centuries of neurology, one century of
psychology, and a half-century of neurobiology and cognitive
neuroscience -- but it can't be truly inconsistent with any of it.
A theory of mind needs a lot of explanatory power, while still
being specific enough to make experimental predictions.
So far, we've actually needed two metaphors: a top-down metaphor that maps thoughts onto ensembles of
neurons, and a bottom-up metaphor that accounts for how
ideas emerge from those apparently chaotic neuron ensembles.
But the neocortical Darwin Machine may well do for both
metaphors -- it might, indeed be more mechanism than
metaphor, thanks to those express-train patterns in the cortex.
Let me turn now to how complex patterns might self-organize, using such Darwinian competitions to embed suitable
resonances in the neural feltwork as we gain experience.
The Search for Hidden Patterns
Our passion for discovering patterns seems to have a lot to
do with our notions of consciousness. If we are to have
meaningful, connected experiences -- ones that we can
comprehend and reason about -- we must be able to discern
patterns to our actions, perceptions, and conceptions. Underlying our vast network of interrelated literal meanings (all of
those words about objects and actions) are those imaginative
structures of understanding such as schema and metaphor,
such as the mental imagery that allows us to extrapolate a path,
or zoom in on one part of the whole, or zoom out until the trees
merge into a forest.
Early childhood contains a number of pattern-finding
challenges, and children seem extraordinarily acquisitive of
ever-more-complex patterns hidden in the sounds and events
that surround them. In our first year of life, we discovered
phonemes within words. After another year of acquiring
words, we were busy discovering schemas and syntax within
word strings, and then we went on to discover nested
embedding and narrative principles among more extended
discourses. The hexagonal superpositions, so like the different
voices of a symphonic performance, show us a way that new
associations can be represented in the brain -- and the
Darwinian aspect suggests how quality could be shaped up via
the usual variation, competition, and inheritance.
When we think seriously as adults, we think even more
abstractly. We conjure up simplified pictures of reality called
concepts or models. We can even discover patterns in
speculative scenarios, as when we create a forwards-leaping
chain of inferences (especially handy for speculating about
consciousness!). As Paul Valéry once said, thought is all about
"that which does not exist, that which is not before me, that
which was, that which will be, that which is possible, that
which is impossible."
Passive awareness (and its neural correlates) may be
much simpler than the creative constructs implied by the
James-Piaget-Popper levels of consciousness; a pop-through
recognition of a familiar object may not need to utilize a
cloning competition with alternatives in the manner of an
ambiguous percept or a novel movement. Hexagonal mosaics
surely aren't everything going on in the brain; indeed, they are
probably just one mode of operation of some expanses of
neocortex, and regulated by other brain regions such as hippocampus and thalamus. But here-this-minute, gone-the-next
mosaics seem quite suitable for explaining many aspects of
mind, aspects that have been difficult to imagine emerging
from quantum mechanics, chemistry, neurotransmitters, single
neurons, simple circuits, or even the smaller neocortical
modules such as minicolumns. In some regions, at some times,
hexagonal competitions might be the main thing happening.
There emerges from this view of our brain, with its relentless
rearrangement from moment to moment, some glimpses of the
neural foundations on which we construct our utterances and
think our thoughts, some possibilities for implementing our
kind of language and rational thought. Dueling choirs are at a
level of explanation that looks as if it might be appropriate for
creativity and decision-making. It will be interesting to see
how much of an explanation for talking-to-yourself
consciousness can be provided by the same quality-bootstrapping process that Charles Darwin discovered back in
1838.
REFERENCES
Derek Bickerton (1990), Language
and Species (University of Chicago
Press).
Jacob Bronowski (1973), The Ascent of
Man (Little, Brown), pp.
348-349.
William H. Calvin (1987), “The brain
as a Darwin Machine,” Nature 330:33-34.
William H. Calvin (1995), “Cortical
columns, modules, and Hebbian cell assemblies,” in Handbook
of Brain Theory and Neural Networks, M. A. Arbib, ed. (MIT Press),
pp. 269-272.
William H. Calvin (1996a). How Brains Think:
Evolving Intelligence, Then and Now (Basic Books, New
York).
William H. Calvin (1996b). The Cerebral
Code: Thinking a Thought in the Mosaics of the Mind (M.I.T. Press,
Cambridge
MA).
William H. Calvin (1997). “The six
essentials? Minimal requirements for the darwinian bootstrapping of
quality,” Journal of Memetics 1, at http://www.fmb.mmu.ac.uk/jom-emit/1997/vol1/calvin_wh.html
Richard Dawkins (1993), “Viruses of
the mind,” in Dennett and His Critics: Demystifying Mind,
edited by Bo Dahlbom
(Blackwell).
Daniel C. Dennett (1991), Consciousness
Explained (Little,
Brown).
Douglas R. Hofstadter (1985), Metamagical
Themas (Basic Books), p.
787.
John H. Holland (1992), “Genetic
algorithms,” Scientific American 267(1):66-72.
Mari Jibu and Kunio Yasue (1995), Quantum
Brain Dynamics and Consciousness: An Introduction. John
Benjamins,
Amsterdam.
Jennifer S. Lund, Takashi Yoshioka,
Jonathan B. Levitt (1993), “Comparison of intrinsic
connectivity in different areas of macaque monkey cerebral
cortex,” Cerebral Cortex 3:148-162.
John Maynard Smith and Eörs Szathmáry
(1995), The Major Transitions of Evolution (Freeman).
Barbara A. McGuire, Charles D. Gilbert,
Patricia K. Rivlin, Torsten N. Wiesel (1991), “Targets of
horizontal connections in macaque primary visual cortex,”
Journal of Comparative Neurology 305:370-392.
Heinz Pagels, The Dreams of Reason (Simon &
Schuster
1988).
Roger Penrose (1994), Shadows of the Mind: A
Search for the Missing Science of Consciousness (Oxford University
Press).
Alan Peters, E. Yilmaz (1993), Neuronal
organization in area 17 of cat visual cortex. Cerebral Cortex 3:49-68.
Karl Popper (1979), quoted by Raphael Sassower in
Cultural Collisions: Postmodern Technoscience
(Routledge).
Hugh Smith (1935),“Synchronous flashing
of fireflies,” Science
82:51.
David Somers and Nancy Kopell (1993),
“Rapid synchronization through fast threshold
modulation,” Biological Cybernetics 68:393-407.
Paul Valéry (1962). quoted at p. 346 in W. H.
Auden and L. Kronenberger, The Viking Book of Aphorisms
(Viking).
Arthur T. Winfree (1967). “Biological rhythms and the behavior of populations of coupled oscillators,” Journal of Theoretical Biology 16:15-42 (1967)
|