Enhancement of Opioid-Mediated Analgesia:
A Solution to the Enigma of Placentophagia
MARK B. KRISTAL
Department of Psychology, State University of New York at Buffalo,
Buffalo, NY 14260
_____
KRISTAL, M.B. Enhancement of opioid-mediated analgesia: A solution
to the enigma of placentophagia. NEUROSCI BIOBEHAV REV 15(3) 425-435, 1991.
-- Two major consequences of placentophagia, the ingestion of afterbirth
materials that occurs usually during mammalian parturition, have been uncovered
in the past several years. The first is that increased contact, associated
with ingesting placenta and amniotic fluid from the surface of the young,
causes an accelerated onset of maternal behavior toward those young. The
second, which probably has importance for a broader range of mammalian
taxa than the first, is that ingestion of afterbirth materials produces
enhancement of ongoing opioid-mediated analgesia. The active substance
in placenta and amniotic fluid has been named POEF, for Placental Opioid-Enhancing
Factor. Recent research on both consequences is summarized, with particular
attention to POEF, the generalizability of the enhancement phenomenon,
its locus and mode of action, and its significance for new approaches to
the management of pain and addiction.
|
Placenta
|
Placentophagia
|
Amniotic fluid
|
Parturition
|
Labor
|
Delivery
|
|
Analgesia
|
Opioids
|
Morphine
|
Addiction
|
Tolerance
|
Withdrawal
|
|
Maternal behavior
|
Mammals
|
POEF
|
Afterbirth
|
Pregnancy
|
Pain
|
PLACENTOPHAGIA, the term generally applied to ingestion of birth membranes
and fluids, is a conspicuous feature of the delivery process in nearly
all nonaquatic placental mammals, whether carnivorous or herbivorous, nesting
or nonnesting, arboreal or terrestrial, monotocous or polytocous, primiparous
or multiparous. No one who has carefully observed parturitional behavior
in mammals in the laboratory or field, in pets, or even in zoo mammals,
can doubt the avidity shown by the mothers in ingesting the afterbirth
(43). In fact, rat mothers give up their newly-delivered pups to the intruding
researcher more readily than they do the placentas (43, 52).
The literature prior to 1980 was fraught with untested speculations
and unverified anecdotes about the proximal causes and the benefits of
placentophagia. Among the causal hypotheses in the literature (which often
did not distinguish between proximal and ultimate causality) were speculations
invoking a general temporary shift to "voracious" carnivorousness; an attempt
to keep the nest site clean to avoid attracting predators; general hunger
resulting from prolonged discomfort and hypophagia during labor; a specific
hunger based on the need to replenish substances (e.g., nutrients) lost
during pregnancy or delivery; and a specific hunger for substances (e.g.,
hormones) that subsequently facilitate lactation or maternal behavior.
In a review paper in 1980 (43), I attempted to organize the research that
had actually been done on the subject, and to dispel some misconceptions.
Among the findings summarized in that paper were the following:
1) Placentophagia seems more properly conceptualized as an ingestive
behavior that is characteristic of maternal females than as a maternal
behavior, in the sense of a caretaking behavior directed toward the neonate,
although elements of both classes of behavior are present. It can be eliminated
with flavor-aversion conditioning procedures (18), and in the absence of
previous experience can be eliminated with lateral hypothalamic lesions
(42). Also, placentophagia (and nest building) can be dissociated from
pup-directed caretaking behaviors at delivery with periventricular medial
preoptic lesions (64). This latter finding was confirmed in a more recent
study on the effects of knife cuts in the hypothalamus on maternal behavior
in rats (23).
2) Placentophagia at delivery does not represent a general shift to
carnivorousness in the monkey (76) or even in an omnivore like the rat
(42), since other meats are refused at that time.
3) Delivery in the rat is not preceded by a period of hypophagia (54),
which suggests that at least in the rat, general hunger is not the explanation
of parturitional placentophagia.
4) Placentophagia at delivery in rats, except perhaps during adverse
conditions, is not critical for normal maternal behavior, lactation, or
postpartum estrus (18, 29, 30, 70) (although the authors of one obscure
and ambiguous report of a human experiment have suggested that there is
a beneficial effect on lactation of postnatal ingestion of freeze-dried
human placenta [71]). It should be noted, however, that all the early studies
examined the effects ingestion of placenta per se, not of amniotic fluid.
5) Nulliparous nonpregnant rats and mice show a dichotomous response
to donor placenta: either they eat it immediately and in great quantity,
or they react as if they are afraid of it, and attempt to escape from it
(43, 46, 56, 63). Although about 40% of the virgin rats that are initially
repelled by donor placenta become enthusiastic eaters during the last week
of pregnancy, the same proportion of nonpregnant rats become placentophagic
after other types of "stressful" events. The major shift toward the avid
placentophagia that is characteristic of the delivery period (virtually
100% eat most, if not all, the placentas delivered) is not observed until
only a few hours before delivery in normal rats (47). The tendency of pregnant
females to become attracted to afterbirth materials only in the last few
hours before delivery was documented subsequently in ewes (59). That a
substantial proportion of virgin rats are attracted to placenta and ingest
it enthusiastically argues against the notion that placentophagia is caused
primarily by a specific hunger for substances depleted during pregnancy
or delivery.
6) There seem not to be any human cultures on record that routinely
practice, or practiced, placentophagia. Placentophagia was defined, however,
as ingestion of the placenta, in particular; ingestion of amniotic fluid
during delivery was not considered. (More on this later.)
BIRTH MATERIALS AND ATTRACTIVENESS OF NEONATES
Some theorists have speculated that different taxonomic groups of mammals
derive different benefits from placentophagia (32). In contrast, our approach
has been to search for one or a few general consequences that could serve
as an adaptive advantage to a wide range of taxa. However, attempts to
document beneficial consequences, either to mother or young, of placentophagia
at parturition were at first frustrating. Initially, an immunological benefit
was hypothesized: it was suggested that placentophagia might reduce the
tendency of the mother's system to produce antibodies to the offspring's
paternal antigens, i.e., to become immune to those antigens as a consequence
of the first parturition, as in erythroblastosis fetalis (43). The possibility
was also suggested that placentophagia stimulated the mother to develop
antibodies against placental tissue remaining in utero; the resulting
immunity (leading to tissue rejection) might, in turn, reduce the possibility
of choriocarcinoma (43). To date these hypotheses have been neither confirmed
nor rejected.
The first important benefit of placentophagia that we found related
to an idea originally suggested by Birch (5): ingestion of placenta and
amniotic fluid on the skin of rat pups by virgin female rats screened for
spontaneous attraction to placenta, increased contact between the adult
and pups, and hastened the induction of maternal behavior toward those
pups (55). This facilitation of the onset of maternal behavior by placenta
and amniotic fluid on the neonate has also been documented in dogs (17)
and in sheep (57, 58).
We asked two questions about this benefit of placentophagia. The first
was whether afterbirth materials on the neonate function simply as powerful
contact-generating attractants no different in kind from other attractants
(55, 63) that also hasten the appearance of maternal behavior. [In our
1981 paper (55), we reported that pups smeared with rat liver or with a
mash of chocolate-chip cookies and milk elicited maternal behavior more
rapidly than a control group comprising pups smeared with saline, saccharin,
or nothing.] If ingestion of afterbirth materials has some effect in addition
to its contact-promoting one, then their ingestion in circumstances that
do not generate such contact should nevertheless hasten the onset of maternal
behavior. However, what we found was that placenta eaten from a dish placed
next to untreated pups did not shorten the latency for virgins to become
maternal toward the pups. We concluded, therefore, that placenta and birth
fluids do not contain qualitatively special attractants or maternal-behavior
inducers, but rather that any material on the skin of the pups that the
adult female found extremely attractive would facilitate the onset of maternal
behavior toward those pups by inducing proximity and contact, thereby providing
the virgin with more stimulation from the pups themselves (55). Terkel
and Rosenblatt had demonstrated a similar effect on the rapidity of induction
of maternal behavior in rats by reducing the size of cages in which pups
and a virgin adult female were co-housed (74).
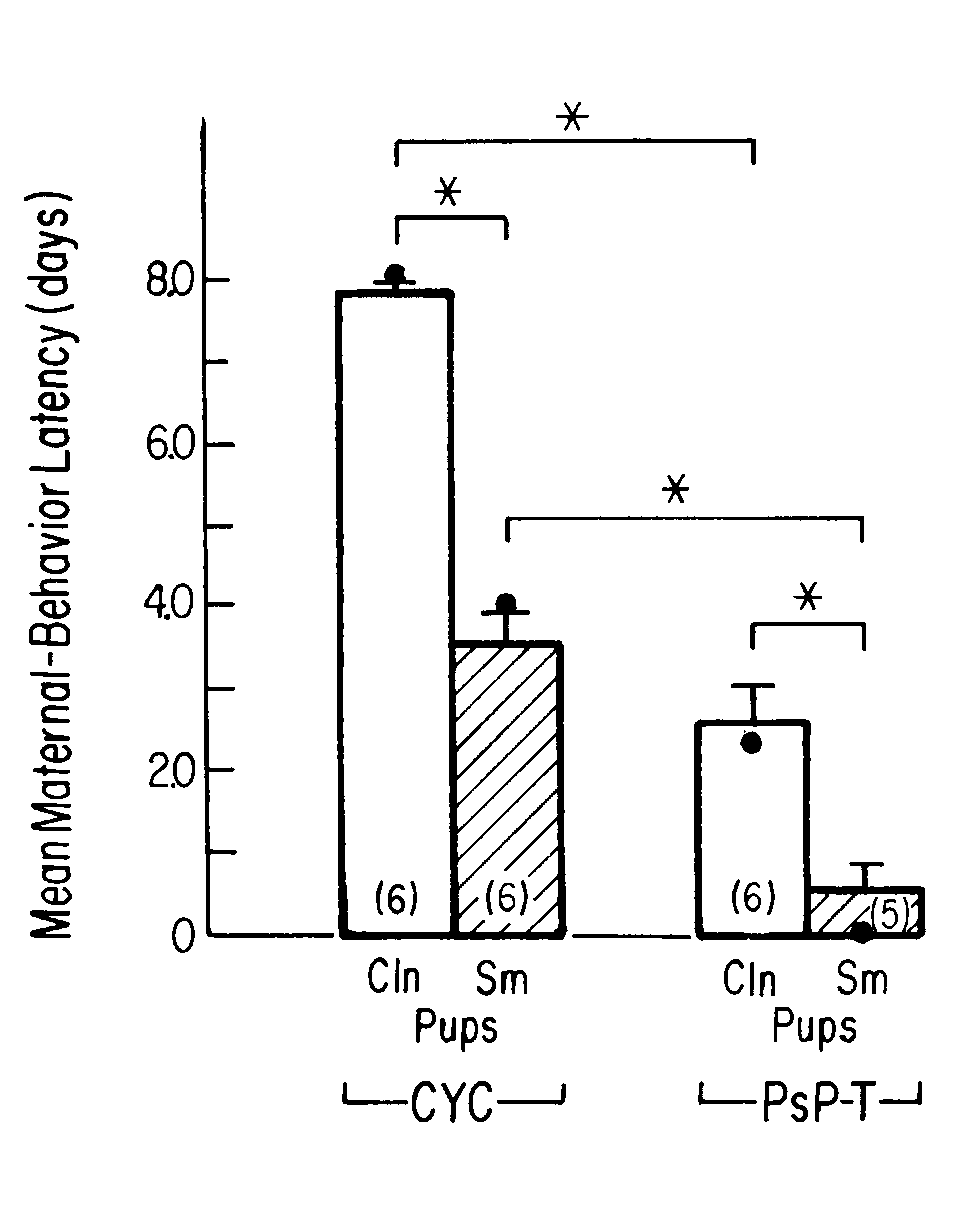
FIG. 1. Mean number of days (±
S.E.M.) to the onset of maternal behavior in rats showing estrous cyclicity
(CYC) or undergoing natural termination of pseudopregnancy (PsP-T) that
were presented with either clean (Cln) pups or pups smeared with placenta
and amniotic fluid (Sm). (Number in parentheses = n; ·
= median. * = p < 0.05.) [from (73) with permission. Ó
1987 APA].
The second question was whether the hormonal state of parturient
rats and birth materials on the skin of pups were additive factors in inducing
the onset of maternal behavior. We approached this question using a pseudopregnancy-termination
model (73), because the hormonal changes occurring during the termination
of pseudopregnancy are qualitatively very similar to those occurring during
the natural termination of pregnancy (31). We found that rats with terminating
pseudopregnancy that were presented with placenta-smeared pups were maternal
sooner than were either cycling rats presented with placenta-smeared pups
or terminating-pseudopregnancy rats presented with clean pups (see Fig.
1). We concluded that the hormonal state of the adult female at delivery
increases the salience and attractiveness of the stimuli emanating from
the pups, and from amniotic fluid and placenta, and helps to bring about
almost immediate pup-directed maternal behavior at that time (73).
A study conducted in sheep had also addressed the issue of attractiveness
to amniotic fluid on neonates and the emergence of maternal behavior at
parturition (57). Washed lambs were not mothered as readily as unwashed
lambs. Interpretation of this result is however rendered ambiguous by the
possibility that the washing introduced an aversive quality. It might have
been useful for the investigators to have washed all the lambs and then
to have coated some of them with amniotic fluid or other attractive substances.
PLACENTAL OPIOID-ENHANCING FACTOR (POEF): THE BASIC PHENOMENON
The second major benefit of placentophagia we have been investigating
concerns the effect of placenta ingestion on opioid-mediated processes,
and was first documented in a study we conducted in 1985. The impetus for
this study derived from a general interest that had been building up on
the part of many researchers, beginning in about 1980, in the role of endogenous
opioids and changes in pain threshold and other opioid-mediated phenomena
during pregnancy and parturition. Although not unequivocal, evidence from
both rat and human studies suggested that plasma and CNS levels of endogenous
opioids rise during pregnancy and peak at or near delivery (19, 21, 28,
33, 40, 66, 67, 78). Further, pain threshold rises at the end of pregnancy,
peaks at or around delivery, and returns to the nonpregnant level by 9-12
hours postpartum (3, 25, 27); Gintzler has dubbed this naltrexone-reversible
change in pain threshold "pregnancy-induced analgesia". Although the findings,
in rats, of Gintzler and his colleagues, have sparked some controversy
(4, 12), our own research has confirmed them (51), and other research has
extended the concept of "pregnancy-induced analgesia" to humans (10) and
to pseudopregnant rats (26).
Our initial idea was that ingestion of placenta during delivery contributed
to the return of pain thresholds to the lower, nonpregnant levels, which
were reinstated by 9-12 hours after delivery. To test this possibility,
we conducted a pilot study in which nonpregnant rats, with pain thresholds
elevated by morphine injection, were fed donor rat placenta. We anticipated
that after ingestion of donor placenta, pain threshold, measured as tail-flick
latency, would fall more rapidly than in rats fed a control meat (ground
beef). The results were quite the reverse of what we had anticipated: the
morphine-injected rats ingesting rat placenta had significantly higher
pain thresholds than did those ingesting ground beef, and the morphine
analgesia lasted longer in the placenta-fed rats than it did in the beef-fed
rats. We recognized that enhanced analgesia might be a far more important
consequence of placentophagia than the one we had originally hypothesized,
and immediately set about designing a formal study in which to begin testing
a new hypothesis, namely, that placentophagia is involved in the elevation
of pain threshold.
The first published study showing an enhancing effect of placenta ingestion
on opioid-mediated analgesia (52) demonstrated that (a) nonpregnant rats
ingesting donor placenta showed significantly more analgesia after intraperitoneal
injection of a threshold dose (3 mg/kg) of morphine sulfate (see Fig. 2),
or after inescapable footshock (2.5 mA for 90 or 120 sec), than did nonpregnant
rats ingesting ground beef, or nothing. However, (b) placenta ingestion
did not produce analgesia in rats not receiving morphine or footshock.
(c) Treatment with the opiate antagonist naltrexone blocked the opioid
component of hindpaw-shock-induced analgesia (79) and also rendered the
treated rats refractory to the enhancing effect of ingested placenta. Finally,
(d) small amounts of placenta enhanced analgesia whereas large amounts
did not. We concluded that whereas ingestion of donor placenta does not
produce analgesia in nonpregnant female rats, ingestion of small amounts
enhances existing opioid-mediated analgesia.
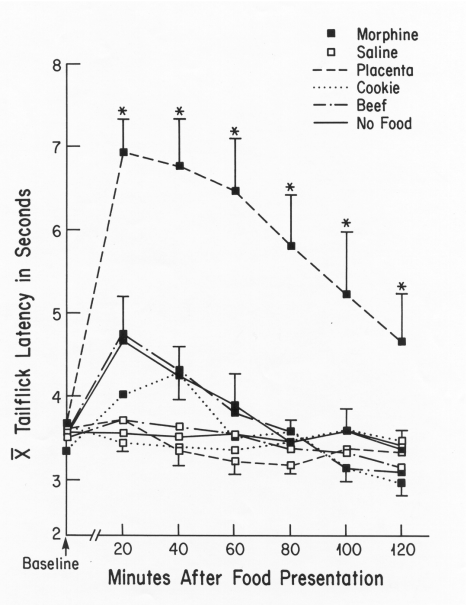
FIG. 2. Mean tail-flick latencies (±
S.E.M.) of virgin rats that received either a morphine (threshold dose,
3 mg/kg) or saline injection, then a 20-min exposure to either placenta,
cookie mash, ground beef, or an empty dish. The Morphine + Placenta Group,
which had the same mean baseline latency as the other groups, showed significantly
longer latencies than all other groups on all trials (*p < 0.05).
[from (52) with permission].
Our next step was to confirm the enhancement of opioid-mediated
analgesia by placenta ingestion in another laboratory, using a different
strain of rat and yet another type of opioid-mediated analgesia. Komisaruk
invited us to attempt the replication in his laboratory, where we could
use his paradigm of vaginal/cervical stimulation-induced analgesia, the
powerful, reliable, stimulus-bound, partly-opioid-mediated form of analgesia
he had been studying extensively (11, 41, 72). That experiment did confirm
our initial finding of enhancement of morphine-mediated analgesia with
placenta ingestion, and also extended the phenomenon to include enhancement
of analgesia produced by vaginal/cervical probing (53). As before, we found
that ingestion of placenta in the absence of ongoing analgesia did not
produce analgesia. It is important to note that mechanical stimulation
of the receptor field of the hypogastric nerve, which is a characteristic
of vaginal/cervical stimulation-induced analgesia, is also characteristic
of the mechanical processes occurring during descent and expulsion of the
fetus during delivery (27, 65). The finding that analgesia produced by
vaginal/cervical stimulation is enhanced by ingestion of placenta was the
first piece of evidence linking placentophagia to analgesia enhancement
in the periparturitional period.
These results precipitated many questions that would have to be addressed.
Among the more pressing were: If placenta is often ingested long after
delivery of the neonate, what is the significance of our finding to events
occurring before and during delivery? What are the dose-response characteristics?
Is the effect of placenta ingestion on analgesia limited to opioid-mediated
analgesia? Is it limited to female rats? Is it limited to rats? Is the
enhancing effect specific for certain types of opioid-mediated analgesia?
Is the effect specific to the tail-flick-latency pain assay? What substance
in placenta is responsible for enhancement? Are opioid-mediated processes
other than analgesia affected by the enhancing substance? What is the mechanism
for enhancement?
Placenta vs. Amniotic Fluid
One aspect of the logic of our hypothesis, that placentophagia elevates
periparturitional pain threshold, i.e., enhances pregnancy-induced analgesia,
remained perplexing. The snag was that the placenta is always delivered
after the fetus in normal deliveries. Therefore, any enhancement
of pregnancy-induced analgesia derived from placentophagia would not affect
pain threshold until after delivery of the first neonate in polytocous
species, and not until the postpartum period in monotocous species. However,
it occurred to us that if ingestion of amniotic fluid also enhances opioid-mediated
analgesia, the hypothesis would still be tenable; amniotic fluid is always
available to the mother during or shortly before the delivery
of a neonate. We tested the effect of amniotic fluid ingestion on morphine-induced
analgesia in a series of studies that allowed nonpregnant rats to eat amniotic-fluid-soaked
ground beef or receive amniotic fluid as an orogastric infusion (49). In
both cases, the amniotic fluid ingestion significantly elevated morphine-induced
analgesia (see Fig. 3); in fact, the effect of amniotic fluid ingestion
was more dramatic than that of placenta ingestion.
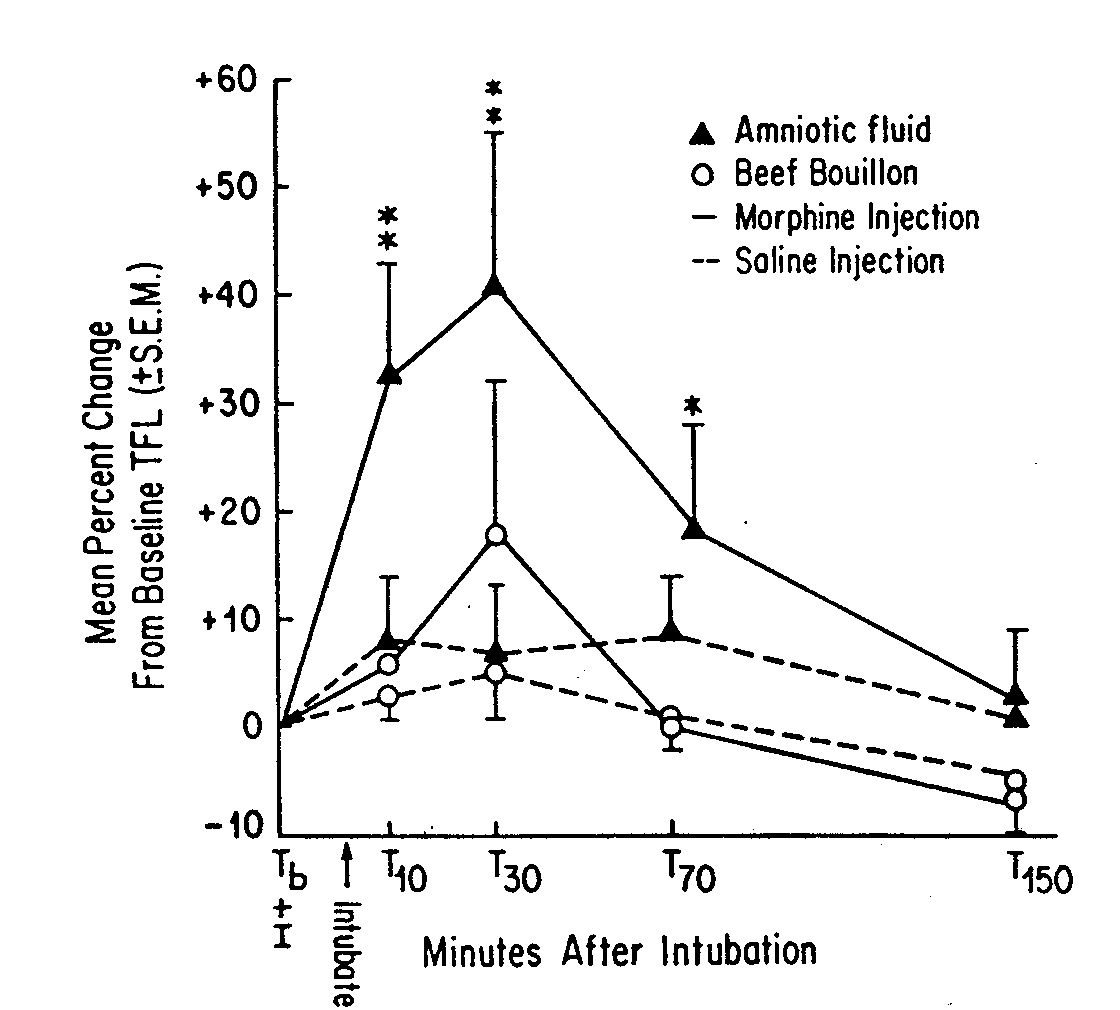
FIG 3. Mean percent change from baseline (±
S.E.M.) tail-flick latency (TFL) of virgin female rats that received either
amniotic fluid or beef bouillon by orogastric tube after receiving an injection
of either morphine (3 mg/kg, IP) or saline (1 ml/kg, IP) (n = 8/group).
I = injection; **significantly different from all other groups, p
< 0.01; *significantly different from the morphine/beef bouillon group,
p < 0.01. [from (49) with permission].
Furthermore, we confirmed that actual ingestion, and not merely
seeing and smelling amniotic fluid, was necessary to produce the enhancement
of analgesia.
At this point we began referring to the putative enhancing substance
as POEF, for placental opioid-enhancing factor.
Dose and Time Course
The next step in developing the case that placentophagia, i.e., ingestion
of either placenta or amniotic fluid, serves to enhance periparturitional
analgesia was to examine the dose-response relationship and time course
of the effect, to see whether these were within the range of parameters
imposed by the delivery process. The dose-response characteristics of placenta
and amniotic fluid were tested first (44). In this series of studies, on
nonpregnant rats, we found that the optimal dose of placenta for enhancement
of a 3-mg/kg (threshold) dose of morphine was about one whole placenta
(500 mg), with doses of 0.25, 0.50, 2 and 4 placentas also producing measurable
enhancement. The optimal dose of amniotic fluid (delivered by orogastric
infusion) for enhancement of the same dose of injected morphine was 0.25
ml, with 0.50 and 1.00 also producing measurable enhancement. Therefore,
at least in regard to enhancement of analgesia produced by injection of
3 mg/kg morphine, the optimal doses of placenta (one) and of amniotic fluid
(0.25 ml), are approximately the amounts that we have found are available
to the rat mother with delivery of each neonate.
We have recently completed a study on the effect of a given dose of
amniotic fluid (0.25 ml) on enhancing the analgesia produced by various
doses of vaginal/cervical stimulation, i.e., produced by various amounts
of pressure applied to the vaginal cervix. In nonpregnant rats in diestrus,
0.25 ml infused amniotic fluid enhanced the analgesia produced by 125 g
vaginal/cervical pressure, but not that produced by 75, 175, or 225 g of
pressure (75). This finding, coupled with the results of an ongoing study
in our laboratory on the enhancing effects of a given dose of amniotic
fluid on analgesia produced by various doses of morphine, and results from
a study on the effects of amniotic-fluid ingestion on morphine-induced
hyperthermia (1, 45), suggests an interaction between the dose of enhancer
and dose of opioid substrate.
The time course of POEF activity was determined by testing for the enhancing
effect of 0.25 ml orogastrically infused amniotic fluid on vaginal/cervical
stimulation-induced analgesia. Nonpregnant rats were again used so that
we might avoid confounding the effect of POEF with physiological changes
occurring during pregnancy. Vaginal/cervical stimulation-induced analgesia
was used because it not only has an abrupt onset and offset, and is therefore
useful in studies involving time course, but it also probably most accurately
represents the form of opioid-mediated (actually partly opioid-mediated)
analgesia that occurs during delivery. We found that enhancement of this
form of analgesia from a single infusion of amniotic fluid was measurable
within 5 minutes after infusion of amniotic fluid, and persisted for about
30 minutes (15). The normal interval between the delivery of successive
rat pups is 20 to 40 minutes.
Therefore, both the dose characteristics and the time course of the
effect of POEF on analgesia in nonpregnant rats fit nicely with the characteristics
of rat parturition.
Nonopioid Analgesics
Although the analgesia-enhancing effect of ingestion of placenta and
amniotic fluid had been demonstrated on a number of different opioid-mediated,
or partly opioid-mediated, forms of analgesia (morphine injection, vaginal/cervical
stimulation, footshock), and the ability of opioid antagonists to block
the effect had also been demonstrated, we had not yet tested for enhancement
of nonopioid analgesia. We used aspirin-mediated analgesia for this test,
because this form of analgesia, particularly in subjects pretreated with
the long-term opioid blocker naltrexone, is considered to be the result
of purely nonopioid processes (20, 36, 81). The experiment was conducted
using the formalin test rather than the radiant-heat tail-flick latency
test. The former, involving inflammation pain produced by a subcutaneous
injection of formalin, is a more sensitive assay for pain threshold in
tests of nonopioid analgesics (20, 36, 37, 81). Since we used morphine-mediated
analgesia as a control, this paradigm also provided us with the opportunity
of testing the effect of ingestion of amniotic fluid on opioid-mediated
analgesia in an assay other than the radiant-heat tail-flick latency test,
one that involves non-reflexive lifting and licking of an inflamed paw
rather than the reflexive movement of the tail.
The results showed that although morphine-mediated analgesia was enhanced
in the formalin test by ingestion of amniotic fluid, aspirin-mediated analgesia
(in naltrexone-pretreated rats) was not (see Fig. 4). Therefore (a) POEF
is ineffective in modifying analgesia produced by at least one major nonopioid
analgesic; and (b) the enhancing effect of POEF on morphine analgesia is
not specific to either thermal pain mechanisms or to neuroanatomical pain
systems related to the tail (48).
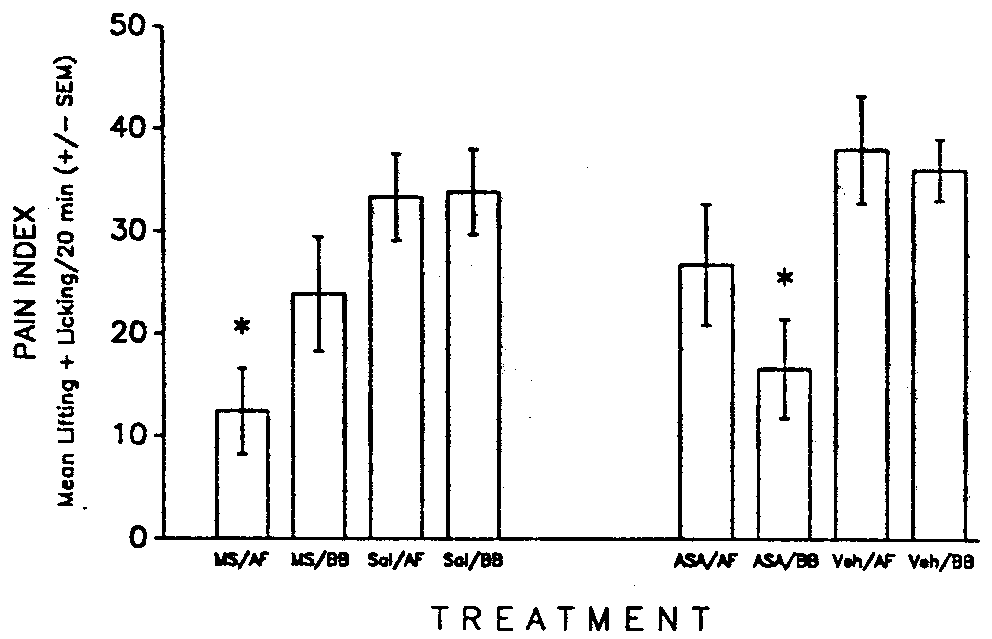
FIG. 4. Pain index (mean s ±
S.E.M. spent paw licking and paw lifting over 20 min) for rats in the morphine
condition (morphine sulfate = MS; saline control = Sal) or the aspirin
condition (aspirin = ASA; vehicle control = Veh) that were infused with
0.25 ml of either amniotic fluid (AF) or beef bouillon (BB). Rats in the
aspirin condition were pretreated with naltrexone. (n = 12/group; *significantly
different from Sal or Veh, p < 0.05). [from (48) with permission].
POEF: ENHANCEMENT OF PERIPARTURITIONAL ANALGESIA
The initial series of studies on the analgesia-enhancing effects of
placenta and amniotic-fluid ingestion was conducted on nonpregnant rats.
The use of nonpregnant rats enabled us to study the basic parameters of
the phenomenon without the confound of ongoing physiological changes occurring
during the rat's 22.5-day pregnancy. However, the use of nonpregnant subjects
prevented us from testing what we considered to be the major benefit of
placentophagia, the enhancement by POEF of pregnancy-mediated analgesia.
A critical test of the effect of parturitional placentophagia on pain
threshold, maternal behavior, lactation, postpartum reproduction, or physiology,
involves an examination of those processes in mothers prevented from ingesting
placenta and amniotic fluid. Methodologically, this group would serve as
the "deprived" group with which normal mothers could be compared. As in
a removal-replacement model, the experimenter could then attempt to eliminate
differences found in the deprived group by returning birth materials to
them for ingestion (the replacement group). However, parturition places
certain limitations on this model. It is extremely difficult (we believe
it actually impossible) to prevent delivering rats from ingesting placenta,
not to mention amniotic fluid, without (a) disrupting parturition, (b)
distressing the mother sufficiently to alter her endogenous opioid levels
and therefore confound the pain-threshold characteristics as well as other
aspects of reproductive physiology (as in [6]), or (c) creating a situation
in which it is technically difficult or impossible to measure pain threshold.
Attempts to remove placentas as they are delivered cause mother rats to
become agitated and to delay or stop delivery. Furthermore, ingestion of
amniotic fluid was found to be at least as effective in the enhancement
of analgesia as was ingestion of placenta. Therefore, since amniotic fluid
is excreted gradually over a relatively long period of time, whereas a
placenta is delivered as a single mass, preventing amniotic-fluid ingestion
presented an even more difficult problem.
The short-term solution to the problem was to examine the effects of
administration of amniotic fluid shortly before the delivering rat's own
amniotic fluid became available for her to ingest (51). Therefore, the
parturient control rats, orogastrically infused with beef bouillon rather
than amniotic fluid, became the "deprived" group, even though they too
were about to give birth and ingest their own amniotic fluid. Some of the
rats were pretreated with naloxone to effect a temporary blockade of opioid
receptors, and thereby reduce pregnancy-induced analgesia. Pain thresholds
were determined using a hot-water tail-withdrawal assay while hand-cradling
the parturient rats. The results showed quite clearly that amniotic-fluid
infusion does enhance pregnancy-induced analgesia. The prepartum pain thresholds
of parturient rats receiving an infusion of donor amniotic fluid 2-6 hours
prepartum were elevated to the level observed during delivery in control
rats, which also ingested their own amniotic fluid during delivery (see
Fig. 5). Additionally, we found that (a) 0.75 ml amniotic fluid produced
more enhancement than did 0.25 ml; and (b) prepartum naloxone treatment
not only eliminated pregnancy-mediated analgesia prior to delivery, but
also rendered the amniotic fluid ineffective in modifying the pain threshold.
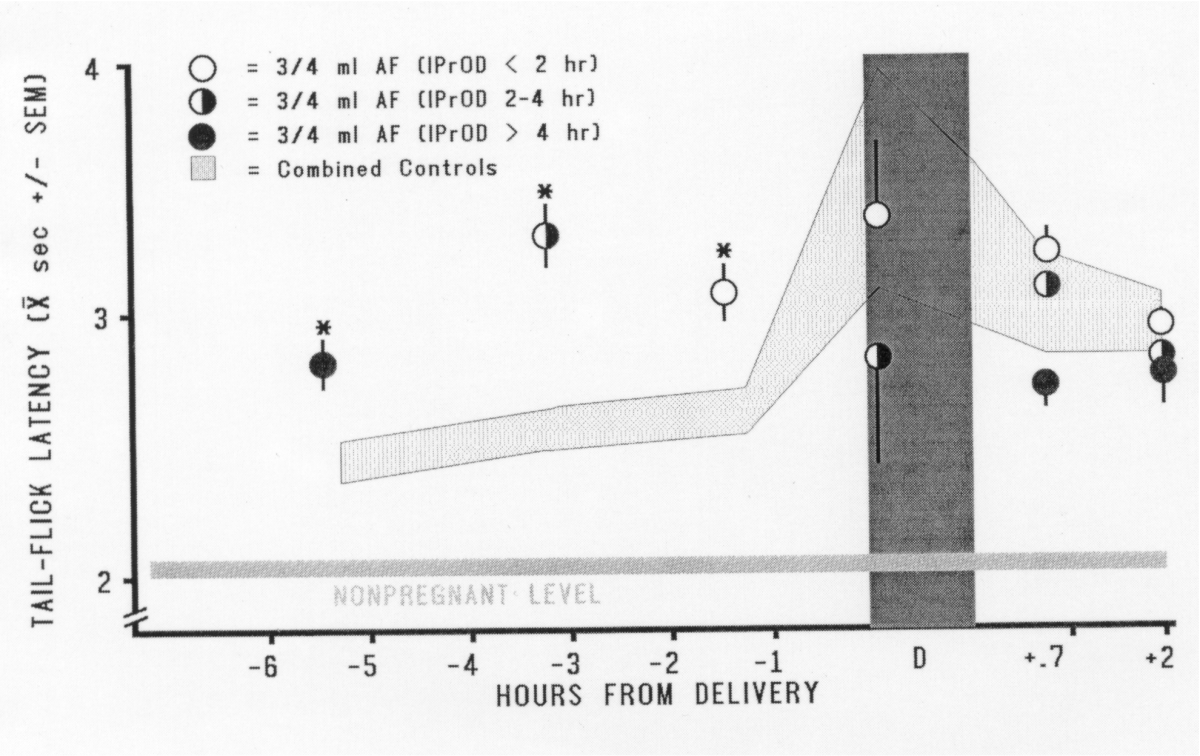
FIG. 5. Effect on pain threshold of infusion of 0.75 ml amniotic
fluid in the hours immediately prior to the onset of delivery. IPrOD =
Interval between the Prepartum TFL test and the Onset
of Delivery. The Combined Controls group consists of controls for
the intubation procedure, for both doses of amniotic-fluid infusion, and
for prepartum testing; these groups did not differ from each other. This
combined group (depicted by the gray band) represents the course of pregnancy-mediated
analgesia. The 0.75-ml amniotic-fluid infusion significantly elevated pregnancy-mediated
analgesia at each of the three IPrODs (*p < 0.01). [from (51)
with permission].
The results were not entirely straightforward. Rats that received
0.25 ml amniotic fluid within two hours before the start of delivery showed
hyperalgesia during the periparturitional period, and many of those showed
heightened aggressiveness for as long as two days afterward. A larger dose
at an interval of two hours or less, or the 0.25-ml dose at longer intervals,
did not produce the same effect.
The effects on other periparturitional phenomena, such as placentophagia,
maternal behavior, and parameters of delivery, were also examined. The
parameters of delivery (onset of delivery after treatment, duration of
delivery, inter-pup interval, pain threshold during and after delivery)
were relatively unaffected either by the amniotic-fluid infusion, which
was administered 1-7 hours prior to the onset of delivery, or by the naloxone
injection, which was administered at least one hour prior to the onset
of delivery. The effects on behavior were more difficult to assess; the
group sizes were large enough for analysis of the pain-threshold data,
but were not sufficient for in-depth analysis of all the behaviors observed.
We did not observe any effects on placentophagia during delivery of either
prepartum naloxone or amniotic-fluid treatment, despite the fact that these
treatments affected pain threshold. However, the dose and time of administration
of naloxone were chosen so as to block opioid receptors prior to the onset
of delivery, but to have little or no effect during delivery, and to be
too low to produce gastrointestinal discomfort (e.g., too low to cause
hypophagia or to be effective in establishing a conditioned flavor aversion).
The main behavioral effects observed in our study on prepartum administration
of amniotic fluid, in addition to the aggression mentioned above, were
the following (50): (a) Rats receiving a low dose (0.25 ml) of amniotic
fluid prior to delivery showed increases in general contractions during
delivery and decreases in general and anogenital self-grooming postpartum.
(b) Rats that received a higher dose (0.75 ml) of amniotic fluid prepartum
showed increases during delivery of the lordotic type of contractions,
in anogenital grooming, and in nest building, all apparently at the expense
of crouching over pups and of resting, which were decreased. (c) The 0.75-ml
prepartum dose of amniotic fluid was associated with postpartum increases
in grouping of pups and decreases in general self-grooming. (d) Some of
the rats receiving 0.75 ml prepartum showed an unusual type of prepartum
contraction involving a hunched body posture, which we called "turtle"
contractions; these were almost never observed in untreated rats or in
rats infused with control fluid. Finally, all these behavioral effects,
observed after prepartum administration of amniotic fluid, were absent
in the amniotic-fluid-treated rats that had been pretreated with naloxone.
More research will need to be done before we can make more general statements
about the role of amniotic-fluid ingestion on periparturitional behaviors.
POEF: GENERALIZABILITY OF EFFECT
How far can the effect of POEF be generalized? As mentioned above, (a)
the effect has been demonstrated to result from ingestion of either rat
placenta or rat amniotic fluid; (b) it is effective in enhancing several
types of opioid-mediated or partially opioid-mediated analgesia but without
effect on nonopioid-mediated analgesia; (c) it has been documented using
several entirely different pain-threshold assays; and (d) enhancement has
been found in both nonpregnant and parturient rats. A recent series of
experiments has been conducted in an attempt to probe further the general
extent of the opioid-enhancement phenomenon.
Males vs. Females
All of the previous studies had been conducted on females, since parturitional
placentophagia is sex-specific. However, male rats, too, are able to experience
the enhancement of morphine-induced analgesia brought about by ingestion
of amniotic fluid (1). Therefore, the mechanism for enhancement is not
sexually dichotomous in rats; it is present in both sexes (see Fig. 6)
and its activation does not require physiological/endocrinological events
associated with female reproductive physiology or with parturition.
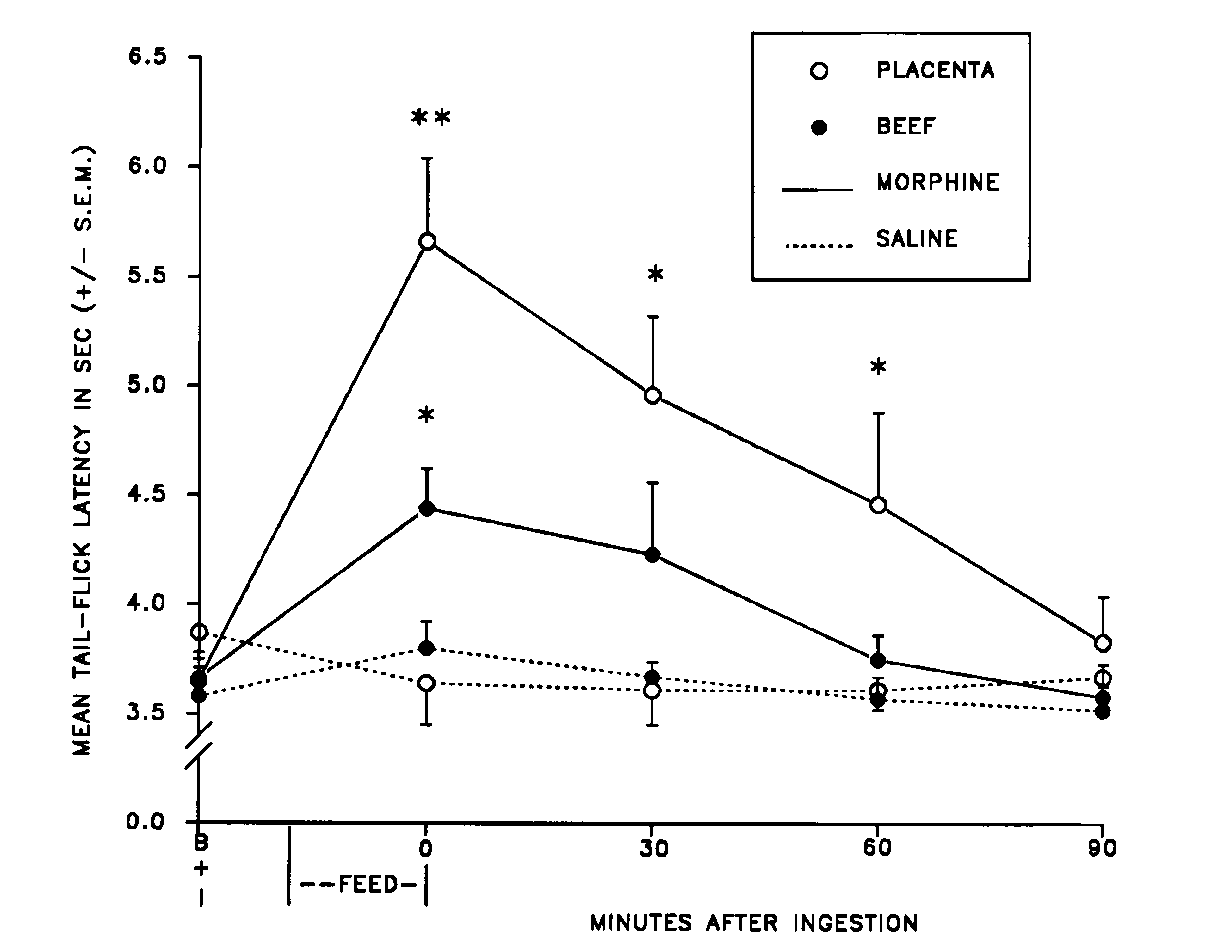
FIG. 6. Mean (± S.E.M.)
tail-flick latency of male rats that ate either placenta or beef after
receiving either a morphine (3 mg/kg, IP) or saline (1 ml/kg, IP) injection
(n = 8/group). I = injection; *significantly different from saline groups
(p < 0.01); **significantly different from all other groups (p
< 0.01). [from (1) with permission].
Species of Donor
Using tail-flick latency in the rat as an assay for POEF activity, we
have documented such activity in homogenates of both human and bottlenose
dolphin (Tursiops truncatus) placenta (1). Both placentas came to us serendipitously:
the human placenta was a healthy, intact, placenta donated by a local mother
and her obstetrician; the dolphin placenta was donated by the Niagara Falls
Aquarium (Niagara Falls, NY). Both placentas were obtained within an hour
or two after delivery and were immediately frozen. Eventually, pieces of
each were thawed and processed. Orogastric infusion of human-placenta cytosol
and voluntary ingestion of small pieces of dolphin placenta were both found
to enhance vaginal/cervical stimulation-induced analgesia in rats. Interestingly,
both humans and dolphins are among the rare exceptions to the mammalian
tendency to engage in parturitional placentophagia regularly (43). These
findings strongly suggest that POEF is a common mammalian substance, which
in turn, suggests the possibility that all mammalian species, including
humans, possess the capacity for responding to POEF.
Route of Administration
Since periparturitional placentophagia is an ingestive behavior, all
our initial studies demonstrating an effect of internalized amniotic fluid
and placenta involved ingestion: substances were either eaten or were infused
orogastrically. To investigate the specific involvement of gastrointestinal
processes, we tested the effect on opioid-mediated analgesia of amniotic
fluid that had been injected either subcutaneously or intraperitoneally
(1). Neither route of injection produced enhancement at the dose of morphine
used, suggesting that gastrointestinal involvement is necessary. This involvement
may take one of a number of forms. One possibility is that the POEF molecule
is activated by the environment of the stomach (involvement of all but
the very beginning of the small intestine is less likely, in light of the
finding that POEF activity can be detected within five minutes after infusion
[15]). Perhaps acidity or enzyme activity is required to form or unbind
the POEF molecule, which is then absorbed into the circulation. An alternative
explanation is that POEF activity depends on contact with receptors found
only in the lining of the stomach (2). Whatever the case, a specific relationship
between an ingestive behavioral process and mediation by specific gastrointestinal
events is not without precedent: the phenomenon of flavor-aversion learning
largely conforms to this model (24), in that flavor is uniquely linkEd
to gastrointestinal discomfort, and not to other forms of discomfort.
POEF in Other Tissue
The animal-derived control substances used in our laboratory (ground
beef, commercial beef bouillon, chicken-egg albumin) have been found not
to contain POEF activity. However, this does not mean that POEF activity
is not to be found in any animal tissues other than afterbirth materials.
To date, the only other substance we have tested that we felt was likely
to possess POEF activity is liver from pregnant rats from which we also
harvested placenta and amniotic fluid. Pregnant-rat liver, when eaten by
nonpregnant rats in amounts equivalent to the optimum effective amount
of placenta (500 mg), did not produce enhancement of morphine-induced analgesia
(1).
Effect on Other Opioid-Mediated Processes -- Thermoregulation
Opioids are involved in phenomena other than analgesia, e.g., feeding,
reproductive behavior, social behavior, and thermoregulation. Differences
may exist between analgesia and these other processes in regard to the
particular opioid receptors involved, whether the process is being mediated
centrally or peripherally, and if centrally, the location within the CNS
of mediation of the processes. To test the generality of the effect of
POEF on opioid-mediated processes, we examined the effect of amniotic-fluid
ingestion on morphine-mediated changes in body temperature (1, 45). Injections
of low doses of morphine (< 5.0 mg/kg) reliably produce hyperthermia
in rats (9, 38, 60). We examined effects of ingestion of amniotic fluid
on both hyperthermia and analgesia in the same rats, in groups receiving
injections of morphine of 0, 2, 3.5 or 5 mg/kg. We found that, as expected,
morphine produced both hyperthermia and analgesia in a dose-dependent fashion,
but only the analgesia was modified by amniotic fluid ingestion (see Fig.
7).
Therefore, POEF probably does not affect all opioid-mediated phenomena.
At this time, we do not know whether differential receptor effects (e.g.,
no enhancement of processes mediated by æ opioid receptors), or different
neuroanatomical loci of action, are primarily responsible for the results.
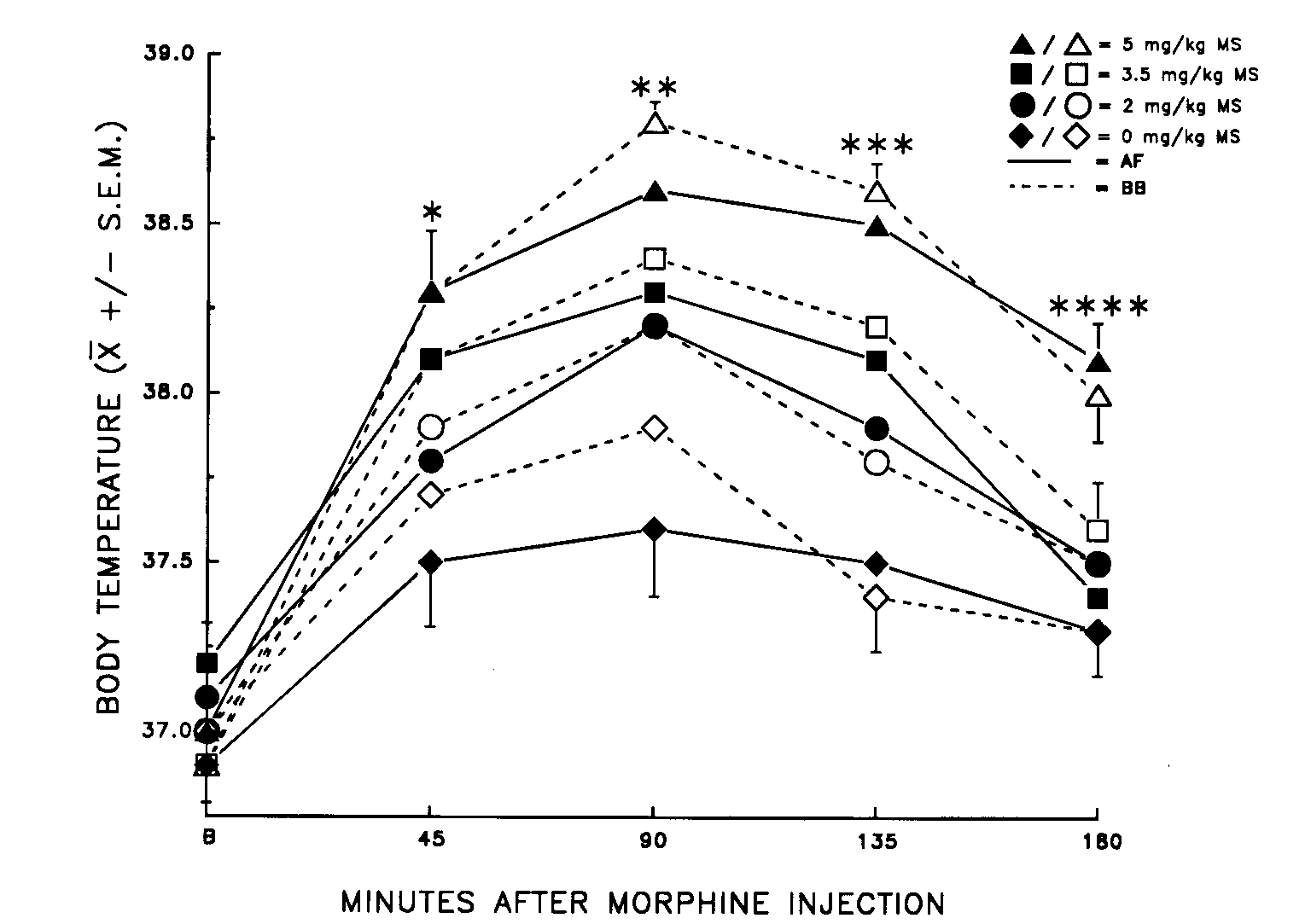
FIG. 7. Mean (±
S.E.M.) body temperature (in ° C) of virgin
female rats before (B) and after injection of morphine (MS) and an orogastric
infusion of amniotic fluid (AF) or beef bouillon control (BB). No significant
differences between AF and BB infusions were found (p > 0.05). Differences
indicated are for morphine dose only (p < 0.05). Pain thresholds
measured at 45 min were significantly higher in the 2 mg/kg MS + AF Group
than in the 2 mg/kg MS + BB Group (p < 0.05). *5 mg/kg = 3.5
mg/kg > 2 mg/kg = 0 mg/kg; **5 mg/kg > 3.5 mg/kg = 2 mg/kg > 0 mg/kg; ***5
mg/kg > 3.5 mg/kg > 2 mg/kg > 0 mg/kg; ****5 mg/kg > 3.5 mg/kg = 2 mg/kg
= 0 mg/kg. [from (1) with permission].
Effect on Other Opioid-Mediated Processes -- Opiate Tolerance
and Withdrawal
Individuals receiving daily injections of a given dose of an opiate
show a diminution of the analgesic response, and many other effects of
opiates, to that dose after several days. Eventually, a stronger dose is
required to elicit the magnitude of response seen initially with the repeated
dose (tolerance). If administration of the opiate is halted abruptly, after
tolerance has been formed, the individual soon manifests a series of transitory
symptoms including hyperalgesia, weight loss, thermoregulatory disturbances,
shakes, and irritability (opiate-withdrawal syndrome) (80). We reasoned
that since POEF enhances morphine-induced analgesia, i.e., reduces the
amount of morphine necessary to produce a given level of analgesia, it
might have a similar effect on the phenomena of morphine tolerance and
withdrawal (16). Rats were made tolerant to a daily injection of 3 mg/kg
morphine. After ten days of treatment, the 3-mg/kg dose no longer produced
analgesia. On Day 11, that dose, coupled with an orogastric infusion of
amniotic fluid, again produced measurable analgesia. A control infusion
of saline on Day 11, to test for dishabituation, did not produce analgesia
in conjunction with 3 mg/kg morphine. The next day (Day 12), the tolerant
rats were allowed to go into withdrawal, during which they exhibited hyperalgesia.
On Day 13, during withdrawal, the rats were treated with 1.5 mg/kg morphine,
or with a vehicle control, and were infused either with amniotic fluid
or with a fluid control. The morphine injection coupled with amniotic-fluid
infusion produced significantly more relief from withdrawal symptoms, particularly
hyperalgesia, than did any other combination of treatments. In addition,
amniotic-fluid infusion in rats receiving no morphine produced a measurable
reduction in withdrawal hyperalgesia (see Fig. 8).
This was the first time we detected an effect of amniotic fluid in the
absence of the induction of opioid-mediated analgesia. It is possible that
POEF enhanced an otherwise undetected endogenous-opioid release occurring
during withdrawal (34).
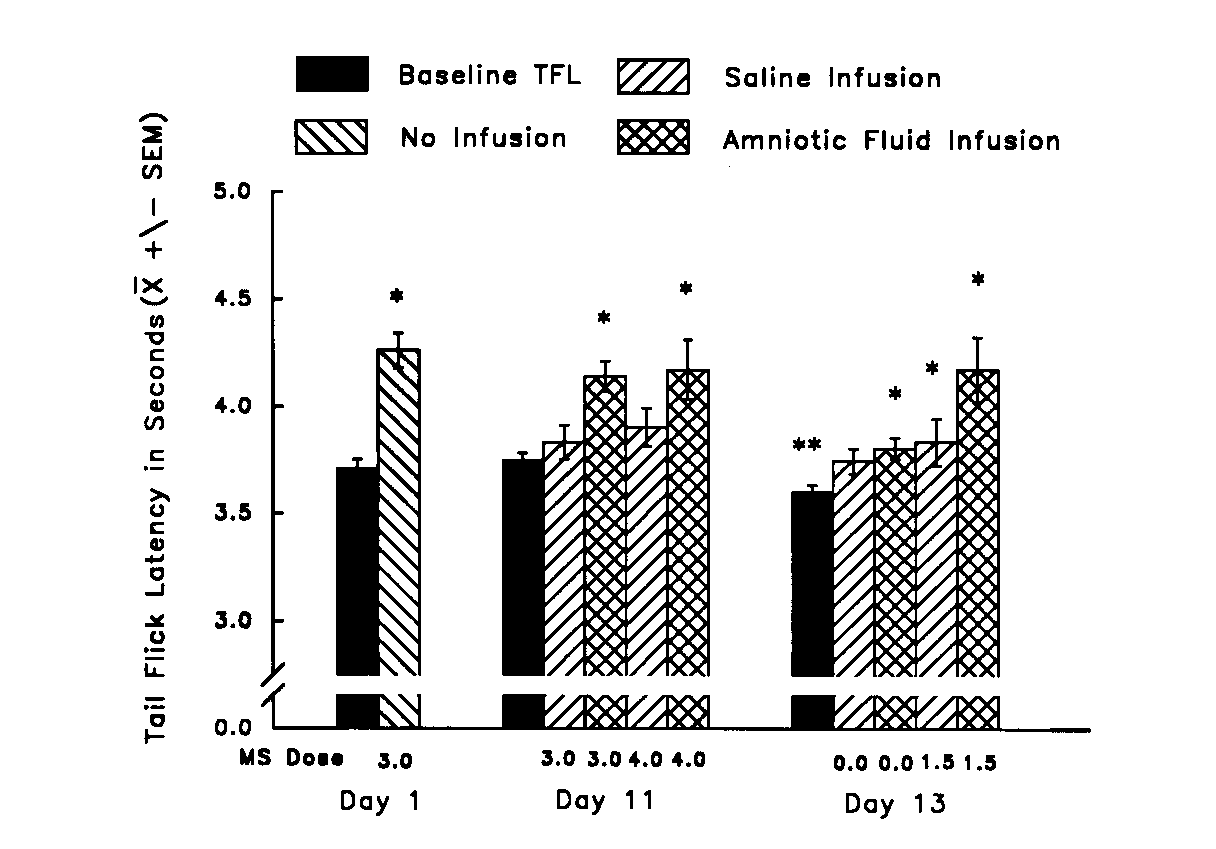
Fig. 8. Mean TFL (±
S.E.M.) of morphine-naive rats (Day 1), morphine-tolerant rats (Day 11),
and rats in morphine withdrawal (Day 13) both before (Baseline) and after
morphine injection and orogastric infusion of either amniotic fluid or
saline. *significantly greater than Baseline on that day (p <
0.05). **significantly lower than Day 1 Baseline (p < 0.01).
[from (16) with permission].
Effect on Other Processes -- Pseudopregnancy
The study we conducted on the enhancing effect of ingestion of 0.25
ml amniotic fluid on analgesia produced by various levels of vaginal/cervical
stimulation produced an unexpected result that we have not yet explored
systematically: amniotic-fluid ingestion tended to block pseudopregnancy
induced by vaginal/cervical stimulation (75). Nonpregnant rats in diestrus
received either 75, 125, 175, or 225 g vaginal/cervical pressure after
receiving an orogastric infusion of 0.25 ml of either amniotic fluid or
saline. Among the rats receiving 225 g vaginal/cervical stimulation, a
significantly greater proportion of those infused with saline entered pseudopregnancy
afterward than did those infused with amniotic fluid. It is not clear yet
what mechanism underlies this effect, or whether POEF or another constituent
of amniotic fluid is responsible. In reducing the likelihood of pseudopregnancy
resulting from vaginal/cervical stimulation, ingestion of amniotic fluid
during delivery may serve also to decrease the probability of pseudopregnancy
resulting from delivery, thereby increasing the chances for the occurrence
of postpartum estrus and consequently a pregnancy eventuating from postpartum
copulation. Although placenta deprivation during parturition was found
to have no effect on the characteristics of postpartum estrus (70), the
effects of deprivation of both placenta and amniotic fluid on the characteristics
of postpartum estrus have not been examined.
MODE AND LOCUS OF POEF ACTION
Perhaps the most burning question is one that if answered first, would
more readily provide answers to the issues of how and where POEF acts:
that question is what is the nature of the POEF molecule? However, until
this question is answered, we have to satisfy ourselves with less direct
approaches to many of the important questions that remain.
Mode of Action of POEF
Virtually all the studies we have conducted on POEF suggest that it
acts by enhancing the action of opioids or opiates that are present in
the system, rather than by producing an increase in endogenous opioids
or because it is, itself, an ingestible opioid. Although placenta and amniotic
fluid are known to contain opioids (22, 35, 39), in our experiments on
non-withdrawing rats administration of amniotic fluid or placenta has never
produced analgesia in rats not treated with an opioid-mediated, or partly
opioid-mediated, analgesic (e.g., 1, 44, 45, 49, 52). If POEF acts by triggering
the release of endogenous opioids, or simply by contributing opioids to
the system, we would have expected to see analgesia resulting from high
levels of intake of placenta or amniotic fluid, whether or not we applied
opioid-mediated analgesia to the subject.
In fact, although our original research suggested that high levels of
intake of placenta or amniotic fluid do not enhance opioid-mediated analgesia
(44), very high doses of POEF may actually be associated with a reduction
of ongoing opioid-mediated analgesia, e.g., hyperalgesia (unpublished observations).
A biphasic response such as this might be explained a number of ways. For
example, the POEF molecule may possess both agonist and antagonist properties,
which emerge differentially at different doses, as is the case with nalorphine,
pentazocine, and cyclazocine (see [61] for review). Alternatively, amniotic
fluid and placenta may contain both opioid enhancers (e.g., POEF) and inhibitors,
but with the inhibitors existing in much smaller concentrations than the
enhancers; when the level of intake of amniotic fluid or placenta in a
brief period of time is extremely high, the dose of inhibitor may approach
optimum whereas the dose of enhancer may be well beyond optimum. Indirect
evidence tends to suggest the first explanation. If POEF does not have
a mixed agonist/antagonist action, then the antagonist molecule in amniotic
fluid must be quite close in size and characteristics to that of POEF,
since the concentration of fractions of amniotic fluid that contain POEF
produces inhibition rather than enhancement of opioid-mediated analgesia
[unpublished observations].
Additional indirect evidence for the mode of action of POEF comes (a)
from the finding that amniotic fluid enhances opioid analgesia when the
fluid is ingested, but not when it is received by subcutaneous or intraperitoneal
injection (1), and (b) from pilot data that show that pretreatment with
infused activated charcoal blocks the enhancing effect of infused amniotic
fluid on opioid-mediated analgesia [unpublished observations]. As mentioned
above, this suggests that either POEF is activated in the gastrointestinal
system and then is absorbed, or it requires binding to and activation of
available gastrointestinal receptors for mediation of the effect. In the
latter case, stimulation of gastric receptors might lead next to either
neural or humoral activity (2). Neural activity, conveyed by vagal afferents,
might ultimately result in direct neural stimulation of brain areas; involvement
of vagal afferent information in the modulation opioid-mediated pain has
already been documented (62, 68). Humoral activity might involve the release
of a second molecule, e.g., cholecystokinin, which might in turn have an
effect on opioid processes.
Locus of Action of POEF
Opioids have both central and peripheral effects. Although the effect
of opioids in the central nervous system is more important for analgesia
than are processes occurring in the periphery, the existence of peripheral
effects makes it necessary to try to separate these from central effects
in order to understand the mode of action of POEF on opioid-mediated analgesia.
Quaternary naltrexone, or naltrexone methobromide, is an opioid antagonist
that, because of its molecular configuration, does not readily cross the
blood-brain barrier. It is therefore quite useful in distinguishing central
from peripheral opioid actions (8, 77). Using either systemic or intracerebroventricular
application of quaternary naltrexone, in conjunction with either systemic
or intracerebroventricular application of morphine sulfate, we have determined
(13, 14) that POEF enhances only the central action of morphine in morphine-induced
analgesia (see Fig. 9).
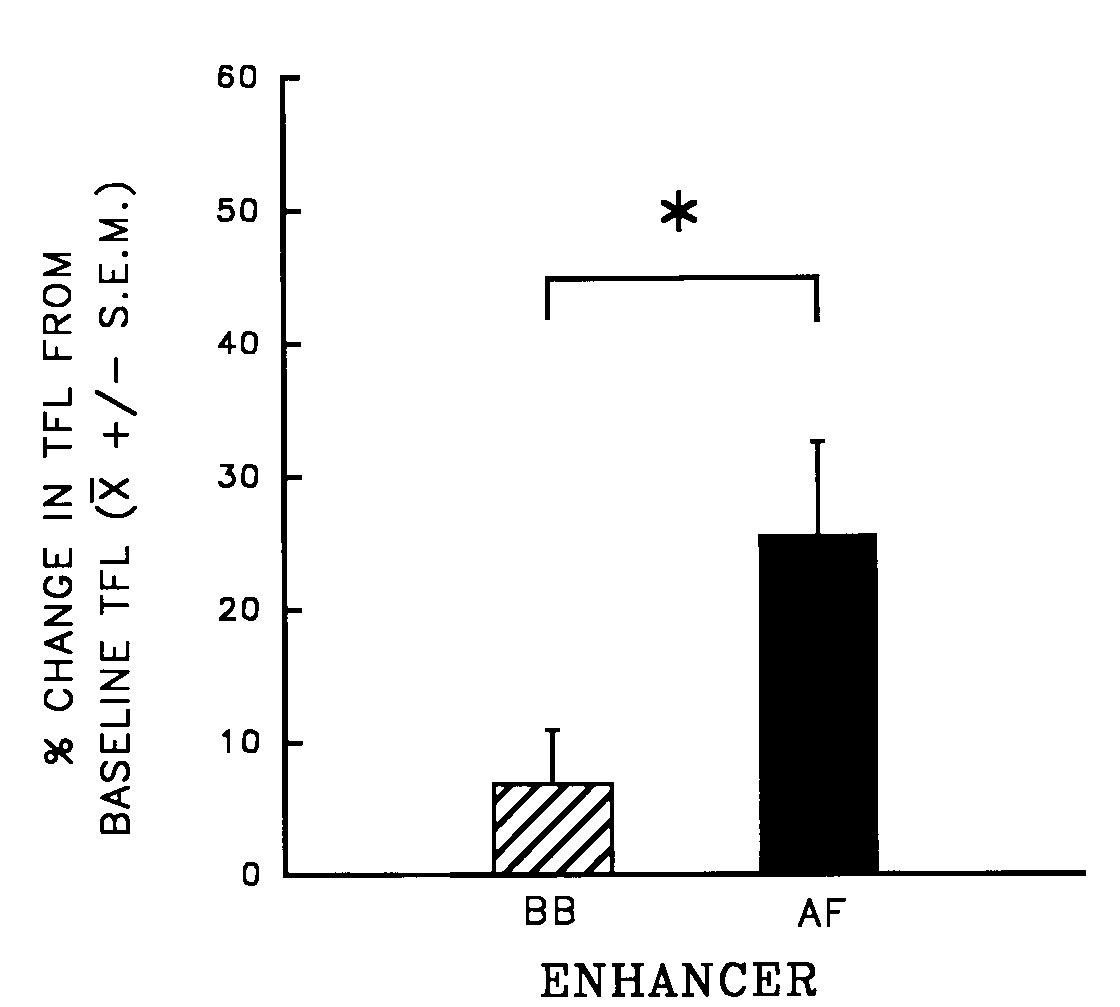
Fig. 9. Mean percent change (±
S.E.M.) from baseline tail-flick latency (TFL) of rats pretreated with
systemic Antagonist [quaternary naltrexone (QN)], then treated with central
Agonist [morphine (MS)] and an orogastric infusion of Enhancer [amniotic
fluid (AF) or beef bouillon control (BB)]. n = 7/group; *p <
0.05. [from (14) with permission].
SUMMARY AND CONCLUSIONS
The important findings of the last decade of research on the consequences
of placentophagia involve two groups of results: those relating to the
effect of placenta and amniotic fluid on the skin of neonates on facilitation
of maternal responsiveness in adults, and those relating to enhancement
of opioid-mediated analgesia.
Studies on the effect on maternal behavior of placenta and amniotic
fluid on the surface of the neonate show that they speed up the rate at
which maternal behavior develops in adults that are not immediately maternal,
and that they probably do so by intensifying the level of adult-infant
contact. In this respect, any manipulation that increases adult-infant
contact, such as applying other attractive substances to the neonate, or
housing the adult and neonate in a very small area, is likely to facilitate
the development of maternal responsiveness on the part of the adult. In
common with mothers of most other mammalian species, rat mothers are intensely
maternal within moments of delivery of the neonate. Therefore, this beneficial
effect of the placenta and amniotic fluid stimuli on the skin of the neonate
is best observed, or may primarily function, in adults in which maternal
responsiveness develops more slowly, such as in nonparturient rats or in
mothers of some nonnesting species.
On the other hand, the effect of ingestion of placenta and amniotic
fluid on enhancement of pregnancy-induced analgesia may be a ubiquitous
mammalian phenomenon, and may have important general consequences both
for mammalian adaptation and, by extrapolation, for the pharmacological
management of pain and of opiate addiction.
Ingestion by the mammalian mother of birth fluids and tissues enhances
the endogenous-opioid-mediated analgesia existing during late pregnancy
and delivery ("pregnancy-induced analgesia"). Amniotic-fluid ingestion
provides enhancement of analgesia early in the delivery process, and placenta
ingestion provides analgesia enhancement later in delivery and could produce
enhancement that lasts into the early postpartum period. The enhancement
seems to be accomplished by potentiating the effect of available opioids,
rather than by increasing opioids either by triggering the release of endogenous
opioids or by adding exogenous opioids to the system. The strategy of producing
more opioid effect without adding more opioids is particularly effective
because too high a level of opioids has been shown to have a deleterious
effect on maternal caretaking behavior (7, 69).
The research on enhancement of analgesia has also shown, among other
things, that (a) the substance responsible for the enhancement (POEF) affects
several types of opioid-mediated analgesia, but not the nonopioid-mediated
analgesia produced by aspirin; (b) POEF ingestion does not, by itself,
produce analgesia; (c) the mechanism for responding to POEF is present
in male as well as nonpregnant female rats; (d) POEF activity can be measured
in a variety of pain assays; (e) although POEF affects tolerance and withdrawal,
in addition to analgesia, it does not affect all opiate-mediated phenomena;
and (f) POEF activity is present in the placenta of mammals other than
rats.
Although we have found that ingestion of amniotic fluid does not modify
morphine-mediated hyperthermia, and therefore does not affect to all opioid-mediated
phenomena, we do not yet know the extent to which it does participate in
opioid-mediated phenomena other than analgesia. We are only now beginning
to understand the possible important effects of ingestion of amniotic fluid
and placenta, and therefore of POEF, on periparturitional behavior and
on postpartum reproductive efficiency.
An interesting question still remains unresolved: why is there so little
evidence that humans groups regularly practice or practiced placentophagia?
An extensive survey of anthropological files revealed no direct evidence
of regular, deliberate, placentophagia among human cultures (43). However,
the focus of the survey, and indeed, the focus of the anthropological notes,
was on the fate of the placenta, not on the fate of amniotic fluid. The
placenta is a large, obtrusive mass of tissue. It easily becomes the focus
of attention during delivery, particularly the attention of observers.
However, in regard to the significance of ingestion of birth materials
and the reduction of periparturitional pain, the placenta may actually
be a red herring. For humans and for many other mammals, the POEF content
of amniotic fluid may be far more important for purposes of periparturitional
pain relief than is that of placenta, although POEF may actually be produced
in the placenta. The availability of amniotic fluid long before expulsion
of the placenta would make it the more useful vehicle for POEF. Parturient
women, in cultures past or present, could have inadvertently ingested amniotic
fluid before expulsion of the fetus perhaps by licking their fingers, or
during delivery by kissing, or cleaning by licking, the amniotic-fluid-soaked
infants; this might be sufficient to produce the effects of ingested amniotic
fluid well before the expulsion of the placenta at the conclusion of delivery.
This form of ingestion of birth materials, and therefore of POEF, was probably
a far more common occurrence than was ingestion of the placenta itself,
but may not have been dramatic enough to warrant attention or documentation.
The more information we gather about POEF, the more likely becomes the
possibility that POEF may provide a significant new direction in pharmacological
management of pain and addiction. For pain management, the strategy of
enhancing the effects of endogenous opioids rather than administering narcotic
analgesics has great appeal for both medical and social reasons. Pain control
by treatment with synthetic POEF may have additional advantages: (a) if
the mechanism of POEF is to potentiate the action of existing opioids rather
than to increase circulating opioids, high doses of POEF are unlikely to
lead to opioid overdose; and (b) if POEF has mixed agonist/antagonist properties,
as some of the research suggests, very high doses -- abusive doses -- may
lead to hyperalgesia. Therapy with POEF may therefore be self-limiting.
Tolerance developed to opiates, and withdrawal from opiates, have also
been found to be modifiable with POEF, in that treatment with POEF reduces
the amount of morphine required to maintain tolerance or to alleviate the
hyperalgesia of withdrawal in morphine-dependent rats. This suggests the
possibility that POEF, when synthesized, may also prove to be a useful
tool in the management and treatment of opiate addiction.
ACKNOWLEDGMENTS
The research on POEF described in this review was supported
primarily by the National Science Foundation (BNS 86-01818 and BNS 88-19837)
and by the National Institute on Drug Abuse (1 R01 DA04586-02). I thank
S. Axelrod, R. J. Barfield, A.R. Caggiula, and B.D. Sachs for their encouragement
and valuable editorial advice on an earlier version of this manuscript.
The American Psychological Association has granted permission for the reprinting
of Figure 1 (copyright 1987). The remaining figures are reprinted with
permission from Pergamon Press.
REFERENCES
-
Abbott, P.; Thompson, A.C.; Ferguson, E.J.; Di Pirro, J.M.;
Doerr, J.C.; Tarapacki, J.A.; Kostyniak, P.J.; Kristal, M.B. Placental
opioid-enhancing factor (POEF): Generalizability of effects. Physiol. Behav.
in press, 1991.
-
Andrews, P.L.R. Vagal afferent innervation of the gastrointestinal
tract. Prog. Brain Res. 67: 65-86; 1986.
-
Baron, S.A.; Gintzler, A.R. Pregnancy-induced analgesia:
effects of adrenalectomy and glucocorticoid replacement. Brain Res. 321:
341-346; 1984.
-
Baron, S.A.; Gintzler, A.R. Effects of hypophysectomy and
dexamethasone treatment on plasma b-endorphin
and pain threshold during pregnancy. Brain Res. 418: 138-145; 1987.
-
Birch, H.G. Sources of order in the maternal behavior of
animals. Am. J. Orthopsychiatry 26: 279-284; 1956.
-
Blank, M.S.; Friesen, H.G. Effects of placentophagy on serum
prolactin and progesterone concentrations in rats after parturition or
superovulation. J. Reprod. Fertil. 60:273-278; 1980.
-
Bridges, R.S.; Grimm, C.T. Reversal of morphine disruption
of maternal behavior by concurrent treatment with the opiate antagonist
naloxone. Science 218: 166-168; 1982.
-
Brown, D.R.; Goldberg, L.I. The use of quaternary narcotic
antagonists in opiate research. Neuropharmacology 24: 181-191; 1985.
-
Clark, W.G. Influence of opioids on central thermoregulatory
mechanisms. Pharmacol. Biochem. Behav. 10: 609-613; 1979.
-
Cogan, R; Spinnato, J.A. Pain and discomfort thresholds in
late pregnancy. Pain 27:63- 68; 1986.
-
Crowley, W.R.; Jacobs, R.; Volpe, J.; Rodriguez-Sierra, J.F.;
Komisaruk, B.R. Analgesic effect of vaginal stimulation in rats: Modulation
by graded stimulus intensity and hormones. Physiol. Behav. 716: 483-488;
1976.
-
Dahl, J.L.; Silva, B.W.; Baker, T.B.; Tiffany, S.T. Endogenous
analgesia in the pregnant rat: An artifact of weight-dependent measures.
Brain Res. 373: 316-323; 1986.
-
DiPirro, J.M.; Thompson, A.C.; Kristal, M.B. Amniotic-fluid
ingestion enhances analgesia produced by the central action of morphine.
Soc. Neurosci. Abstr. 16: 213; 1990.
-
DiPirro, J.M.; Thompson, A.C.; Kristal, M.B. Amniotic-fluid
(POEF) ingestion enhances morphine analgesia centrally but not peripherally.
Physiol. Behav. in press, 1991.
-
Doerr, J.C.; Kristal, M.B. Enhancement of opioid-mediated
analgesia by ingestion of amniotic fluid: Onset latency and duration. Physiol.
Behav. 46: 913-915; 1989.
-
Doerr, J.C.; Kristal, M.B. Effects of amniotic-fluid ingestion
on a characteristic of morphine tolerance and withdrawal in rats. Physiol.
Behav. in press; 1991.
-
Dunbar, I.; Ramson, E.; Buehler, M. Pup retrieval and maternal
attraction to canine amniotic fluids. Behav. Proc. 6: 249-260; 1981.
-
Engwall, D.B.; Kristal, M.B. Placentophagia is modifiable
by taste aversion conditioning. Physiol. Behav. 18: 495-502; 1977.
-
Facchinetti, F.; Centini, G.; Parrini, D.; Petraglia, F.;
D'Antonia, N.; Cosmi, E.V.; Genazzani, A.R. Opioid plasma levels during
labor. Gynecol. Obstet. Invest. 13: 155- 163; 1982.
-
Ferreira, S.H.; Lorenzetti; B.B.; Correa, F.M.A. Central
and peripheral antialgesic action of aspirin-like drugs. Eur. J. Pharmacol.
53: 39-48; 1978.
-
Fletcher, J.E.; Thomas, T.E.; Hill, R.G. An investigation
into opioid systems in the rat. Life Sci. 33: 515-518; 1983.
-
Fraioli, F.; Genazzani, A.R. Human placental beta-endorphin.
Gynecol. Obstet. Invest. 11: 37-44; 1980.
-
Franz, J.R.; Leo, R.J.; Steuer, M.A.; Kristal, M.B. Effects
of hypothalamic knife cuts and experience on maternal behavior in the rat.
Physiol. Behav. 38: 629-640; 1986.
-
Garcia, J.; Hankins, W.G.; Rusiniak, K.W. Behavioral regulation
of the milieu interne in man and rat. Science 185: 824-831; 1974.
-
Gintzler, A.R. Endorphin-mediated increases in pain threshold
during pregnancy. Science 210: 193-195; 1980.
-
Gintzler, A.R.; Bohan, M.C. Pain thresholds are elevated
during pseudopregnancy. Brain Res. 507: 312-316; 1990.
-
Gintzler, A.R.; Peters, L.C.; Komisaruk, B.R. Attenuation
of pregnancy-induced analgesia by hypogastric neurectomy in rats. Brain
Res. 277: 186-188; 1983.
-
Goland, R.S.; Wardlaw, S.L.; Stark, R.I.; Frantz, A.G. Human
plasma beta-endorphin during pregnancy, labor, and delivery. J. Clin. Endocrinol.
Metab. 52: 74-78; 1981.
-
Grota, L.J. Factors influencing the acceptance of caesarean
delivered offspring by the foster mother. Physiol. Behav. 3: 265-269; 1968.
-
Grota, L.J. The effects of placenta and fetal fluids on the
acceptance of foster young. Dev. Psychobiol. 6: 495-502; 1973.
-
Gunnet, J.W.; Freeman, M.E. The mating-induced release of
prolactin: A unique neuroendocrine response. Endocr. Rev. 4: 44-61; 1983.
-
Hart, B.L. The behavior of domestic animals. New York: Freeman;
1985.
-
Hoffman, D.I.; Abboud, T.R.; Haase, H.R.; Hung, T.T.; Goebelsmann,
V. Plasma b-endorphin concentrations prior to
and during pregnancy, in labor, and after delivery. Am. J. Obstet. Gynecol.
150: 492-496; 1984.
-
H"llt, V; Przewlocki, R.; Herz, A. b-Endorphin-like
immunoreactivity in plasma, pituitaries and hypothalamus of rats following
treatment with opiates. Life Sci. 23: 1057-1066; 1978.
-
Houck, J.C.; Kimball, C.; Chang, C.; Pedigo, N.W.; Yamanura,
H.I. Placental b-endorphin-like peptides. Science
207: 78-80; 1980.
-
Hunskaar, S. Similar effects of acetylsalicylic acid and
morphine on immediate responses to acute noxious stimulation. Pharmacol.
Toxicol. 60: 167-170; 1987.
-
Hunskaar, S.; Hole, K. The formalin test in mice: Dissociation
between inflammatory and noninflammatory pain. Pain 30: 103-114; 1987.
-
Jorenby, D.E.; Keesey, R.E.; Baker, T.B. Effects of dose
on effector mechanisms in morphine-induced hyperthermia and poikilothermia.
Psychopharmacology (Berlin) 98: 269- 274; 1989.
-
Kimball, C.D. Do endorphin residues of beta-lipotropin in
hormone reinforce reproductive functions? Am. J. Obstet. Gynecol. 143:
127-130; 1979.
-
Kimball, C.D.; Chang, C.M.; Chapman, M.B. Endogenous opioid
peptides in intrapartum blood. Am. J. Obstet. Gynecol. 149: 79-82; 1984.
-
Komisaruk, B.R.; Wallman, J. Antinociceptive effects of vaginal
stimulation in rats: Neurophysiological and behavioral studies. Brain Res.
137: 85-107; 1977.
-
Kristal, M.B. Effects of lateral hypothalamic lesions on
placentophagia in virgin, primiparous, and multiparous rats. J. Comp. Physiol.
Psychol. 84: 53-62; 1973.
-
Kristal, M.B. Placentophagia: A biobehavioral enigma (or
De gustibus non disputandum est). Neurosci. Biobehav. Rev. 4: 141-150;
1980.
-
Kristal, M.B.; Abbott, P.; Thompson, A.C. Dose-dependent
enhancement of morphine- induced analgesia by ingestion of amniotic fluid
and placenta. Pharmacol. Biochem. Behav. 31:351-356; 1988.
-
Kristal, M.B.; Ferguson, E.J.; Bruschetti, J.; Thompson,
A.C. Placental opioid-enhancing factor does not modify morphine-induced
hyperthermia. Soc. Neurosci. Abstr. 15:845; 1989.
-
Kristal, M.B.; Graber, G.C. Placentophagia in nonpregnant
rats: Influence of estrous cycle stage and birthplace. Physiol. Behav.
17: 599-605; 1976.
-
Kristal, M.B.; Peters, L.C.; Franz, J.R.; Whitney, J.F.;
Nishita, J.K.; Steuer, M.A. The effect of pregnancy and stress on the onset
of placentophagia in Long-Evans rats. Physiol. Behav. 27:591-595; 1981.
-
Kristal, M.B.; Tarapacki, J.A.; Barton, D. Amniotic fluid
ingestion enhances opioid-mediated but not nonopioid-mediated analgesia.
Physiol. Behav. 47: 79-81; 1990.
-
Kristal, M.B.; Thompson, A.C.; Abbott, P. Ingestion of amniotic
fluid enhances opiate analgesia in rats. Physiol. Behav. 38: 809-815; 1986.
-
Kristal, M.B.; Thompson, A.C.; Abbott, P.; Di Pirro, J.M.;
Doerr, J.C.; Ferguson, E.J. Amniotic-fluid ingestion, opioid enhancement
and parturition. Presented at Conference on Reproductive Behavior, Omaha,
NE, June 1988.
-
Kristal, M.B.; Thompson, A.C.; Abbott, P.; Di Pirro, J.M.;
Ferguson, E.J.; Doerr, J.C. Amniotic-fluid ingestion by parturient rats
enhances pregnancy-mediated analgesia. Life Sci. 46: 693-698; 1990.
-
Kristal, M.B.; Thompson, A.C.; Grishkat, H.L. Placenta ingestion
enhances opiate analgesia in rats. Physiol. Behav. 35: 481-486; 1985.
-
Kristal, M.B.; Thompson, A.C.; Heller, S.B.; Komisaruk, B.R.
Placenta ingestion enhances analgesia produced by vaginal/cervical stimulation
in rats. Physiol. Behav. 36: 1017-1020; 1986.
-
Kristal, M.B.; Wampler, R.S. Food and water intake prior
to parturition in the rat. Physiol. Psychol. 1: 297-300; 1973.
-
Kristal, M.B.; Whitney, J.F.; Peters, L.C. Placenta on pups'
skin accelerates onset of maternal behaviour in nonpregnant rats. Anim.
Behav. 29: 81-85; 1981.
-
Kristal, M.B.; Williams, C.L. The effects of strain, reproductive
condition, and strain of placenta donor on placentophagia in nonpregnant
mice. Physiol. Psychol. 1: 354-356; 1973.
-
Levy, F.; Poindron, P. Influence of amniotic fluids in the
manifestation of maternal behavior in parturient ewes. Biol. Behav. 9:
271-278; 1984.
-
Levy, F.; Poindron, P. The importance of amniotic fluids
for the establishment of maternal behaviour in experienced and inexperienced
ewes. Anim. Behav. 35: 1188- 1192; 1987.
-
Levy, F.; Poindron, P.; le Niendre, P. Attraction and repulsion
by amniotic fluids and their olfactory control in the ewe around parturition.
Physiol. Behav. 31: 687-692; 1983.
-
Lotti, V.J. Body temperature responses to morphine. In: The
pharmacology of thermoregulation. San Francisco: Karger; 1972.
-
Martin, W.R. Pharmacology of opioids. Pharmacol. Rev. 35:
283-323; 1983.
-
Ness, T.J.; Gebhart, G.F. Visceral pain -- a review of experimental
studies. Pain 41:167-234; 1990.
-
Nishita, J.K.; Kristal, M.B. Placentophagia in rats: the
attractiveness of placenta. Presented at Eastern Psychological Association,
New York, New York, April 1980.
-
Noonan, M.; Kristal, M.B. Effects of medial preoptic lesions
on placentophagia and on the onset of maternal behavior in the rat. Physiol.
Behav. 22: 1197-1202; 1979.
-
Peters, L.C.; Kristal, M.B.; Komisaruk, B.R. Sensory innervation
of the external and internal genitalia of the female rat. Brain Res. 408:
199-204; 1987.
-
Pilkington, J.W.; Nemeroff, C.B.; Mason, G.A.; Prange, A.J.
Increase in plasma b-endorphin-like immunoreactivity
at parturition in normal women. Am. J. Obstet. Gynecol. 145: 111-113; 1983.
-
Raisanen, I.; Paatero, H.; Salminen, K.; Laatikainen, T.
Pain and plasma b-endorphin level during labor.
Obstet. Gynecol. 64: 783-786; 1984.
-
Ren, K.; Randich, A.; Gebhart, G.F. Vagal afferent modulation
of a nociceptive reflex in rats: Involvement of spinal opioid and monoamine
receptors. Brain Res. 446: 285-294; 1988.
-
Rubin, B.S.; Bridges, R.S. Disruption of ongoing maternal
responsiveness in rats by central administration of morphine sulfate. Brain.
Res. 307: 91-97; 1984.
-
Sachs, B.D. Behavior of rats in the perinatal period. Am.
Zool. 9: 1068; 1969.
-
Soykov -Pachnerov , E.; Brutar, V; Golov , B.; Zvolsk , E.
Placenta as a lactagogon. Gynaecologia 138: 617-627; 1954.
-
Steinman, J.L.; Roberts, L.A.; Komisaruk, B.R. Evidence that
endogenous opiates contribute to the mediation of vaginal-stimulation produced
anti-nociception in rats. Soc. Neurosci. Abstr. 8: 47; 1982.
-
Steuer, M.A.; Thompson, A.C.; Doerr, J.C.; Youakim, M.; Kristal,
M.B. Induction of maternal behavior in rats: Effects of pseudopregnancy
termination and placenta-smeared pups. Behav. Neurosci. 101: 219-227; 1987.
-
Terkel, J.; Rosenblatt, J.S. Aspects of nonhormonal maternal
behavior in the rat. Horm. Behav. 2: 161-171; 1971.
-
Thompson, A.C.; Abbott, P.; Doerr, J.C.; Ferguson, E.J.;
Kristal, M.B. Dose-dependent enhancement of VSIA and blockade of VS-induced
pseudopregnancy by ingestion of amniotic fluid. Physiol. Behav. in press;
1991.
-
Tinklepaugh, O.L.; Hartman, C.G. Behavioral aspects of parturition
in the monkey (Macaca rhesus). J. Comp. Psychol. 11: 63-98; 1930.
-
Valentino, R.J.; Herling, S.; Woods, J.H.; Medzihradsky,
F.; Merz, H. Quaternary naltrexone: Evidence for the central mediation
of discriminative stimulus effects of narcotic agonists and antagonists.
J. Pharmacol. Exp. Ther. 217: 652-659; 1981.
-
Wardlaw, S.L.; Frantz, A.G. Brain b-endorphin
during pregnancy, parturition and the postpartum period. Endocrinology
113: 1664-1668; 1983.
-
Watkins, L.R.; Cobelli, D.A.; Faris, P.; Aceto, M.D.; Mayer,
D.J. Opiate vs non-opiate footshock-induced analgesia (FSIA): The body
region shocked is a critical factor. Brain Res. 242: 299-308; 1982.
-
Wei, E.; Way, E.L. Application of the pellet implantation
technique for the assessment of tolerance and physical dependence in the
rodent. In: Ehrenpreis, S.; Neidle, A., eds. Methods in narcotic research.
New York: Marcel Dekker Press; 1975.
-
Yaksh, T.L. Central and peripheral mechanisms for the antialgesic
action of acetylsalicylic acid. In: Barnett, H.J.M.; Hirsh, J.; Mustard,
J.F., eds. Acetylsalicylic acid: New uses for an old drug. New York: Raven
Press; 1982.